Se acepta ampliamente que el asentamiento de las Américas comenzó cuando los cazadores-recolectores del Paleolítico entraron en América del Norte desde la estepa del mamut del norte de Asia a través del puente terrestre de Beringia , que se había formado entre el noreste de Siberia y el oeste de Alaska debido a la bajada del nivel del mar durante el Último Máximo Glacial . [1] Estas poblaciones se expandieron al sur de la capa de hielo Laurentide y se extendieron rápidamente por América del Norte y del Sur , hace 14.000 años. [2] [3] [4] [5] Las primeras poblaciones de América, antes de hace aproximadamente 10.000 años, se conocen como paleoindios .
El poblamiento de las Américas es una cuestión abierta desde hace mucho tiempo, y aunque los avances en arqueología , geología del Pleistoceno , antropología física y análisis de ADN han arrojado progresivamente más luz sobre el tema, quedan cuestiones importantes sin resolver. [6] Si bien existe un acuerdo general de que las Américas se colonizaron por primera vez desde Asia, el patrón de migración, su momento y el lugar (s) de origen en Eurasia de los pueblos que migraron a las Américas siguen sin estar claros. [3]
Los modelos de migración predominantes describen diferentes marcos de tiempo para la migración asiática desde el estrecho de Bering y la posterior dispersión de la población fundadora por todo el continente. [7] Los pueblos indígenas de las Américas se han relacionado con las poblaciones siberianas por factores lingüísticos , la distribución de los tipos de sangre y la composición genética reflejada por los datos moleculares , como el ADN . [8] [9]
La "primera teoría de Clovis" se refiere a la hipótesis de la década de 1950 de que la cultura Clovis representa la primera presencia humana en las Américas, comenzando hace unos 13.000 años; La evidencia de culturas anteriores a Clovis se ha acumulado desde 2000, lo que hace retroceder la posible fecha del primer poblamiento de las Américas a hace 33.000 años. [10] [11] [12] Además, los huesos de fauna descubiertos en la cueva Coxcatlán , que datan de entre 33.448 y 28.279 años, indicaron que la llegada de humanos a América del Norte podría haber ocurrido antes del período del Último Máximo Glacial. [13]
El medio ambiente durante la última glaciación
Para obtener una introducción a las técnicas de datación por radiocarbono utilizadas por arqueólogos y geólogos, consulte la datación por radiocarbono .
Emergencia y sumersión de Beringia
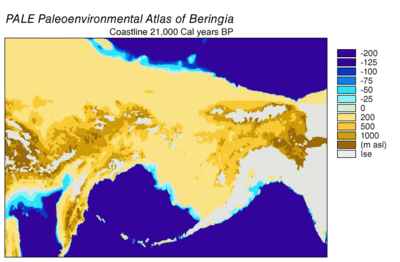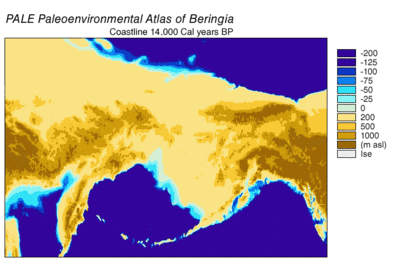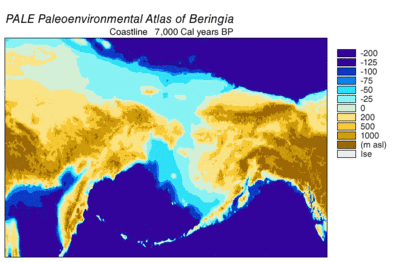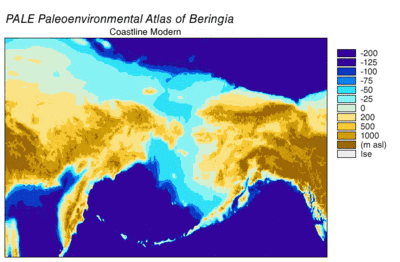
Durante la glaciación de Wisconsin , el agua de los océanos de la Tierra se almacenó, en diversos grados con el tiempo, en el hielo de los glaciares . A medida que el agua se acumuló en los glaciares, el volumen de agua en los océanos disminuyó en consecuencia, lo que resultó en un descenso del nivel global del mar . La variación del nivel del mar a lo largo del tiempo se ha reconstruido mediante el análisis de isótopos de oxígeno de núcleos de aguas profundas, la datación de terrazas marinas y el muestreo de isótopos de oxígeno de alta resolución de cuencas oceánicas y casquetes de hielo modernos. Una caída del nivel del mar eustático de unos 60 a 120 metros (200 a 390 pies) desde los niveles actuales, comenzando alrededor de 30.000 años antes de Cristo , creó Beringia., una característica geográfica extensa y duradera que conecta Siberia con Alaska. [14] Con el aumento del nivel del mar después del Último Máximo Glacial (LGM), el puente terrestre de Beringia quedó nuevamente sumergido. Las estimaciones del resurgimiento final del puente terrestre de Beringia basadas puramente en la batimetría actual del Estrecho de Bering y la curva eustática del nivel del mar sitúan el evento alrededor de 11.000 años AP (Figura 1). La investigación en curso que reconstruye la paleogeografía beringiana durante la desglaciación podría cambiar esa estimación y una posible inmersión temprana podría restringir aún más los modelos de migración humana a América del Norte. [14]
Glaciares
El inicio del Último Máximo Glacial después de 30.000 años AP vio la expansión de los glaciares alpinos y las capas de hielo continentales que bloquearon las rutas de migración fuera de Beringia. Hacia 21.000 años antes de Cristo, y posiblemente miles de años antes, las capas de hielo Cordilleran y Laurentide se fusionaron al este de las Montañas Rocosas , cerrando una posible ruta de migración hacia el centro de América del Norte. [15] [16] [17] Los glaciares alpinos en las cadenas costeras y la península de Alaska aislaron el interior de Beringia de la costa del Pacífico. Los glaciares alpinos costeros y los lóbulos del hielo cordillerano se fusionaron en los glaciares del piedemonteque cubría grandes extensiones de la costa tan al sur como la isla de Vancouver y formó un lóbulo de hielo a través del Estrecho de Juan de Fuca por 15.000 años 14 C AP (18.000 años cal AP). [18] [19] Los glaciares alpinos costeros comenzaron a retroceder alrededor de 19.000 años cal AP [20] mientras que el hielo cordillerano continuó avanzando en las tierras bajas de Puget hasta 14.000 años 14 C AP (16.800 años cal AP). [19] Incluso durante la extensión máxima de hielo costero, persistieron refugios no glaciares en las islas actuales, que albergaron mamíferos terrestres y marinos.[17] A medida que se produjo la desglaciación, los refugios se expandieron hasta que la costa quedó libre de hielo en 15.000 años cal AP. [17] El retroceso de los glaciares en la península de Alaska proporcionó acceso desde Beringia a la costa del Pacífico alrededor de 17.000 años-cal AP. [21] La barrera de hielo entre el interior de Alaska y la costa del Pacífico se rompió a partir de 13.500 años 14 C (16.200 años cal) AP. [18] El corredor libre de hielo hacia el interior de América del Norte se abrió entre 13.000 y 12.000 años cal AP. [15] [16] [17] La glaciación en el este de Siberia durante el LGM se limitó a los glaciares alpinos y de los valles en las cadenas montañosas y no bloqueó el acceso entre Siberia y Beringia. [14]
Ambientes climáticos y biológicos
Los paleoclimas y la vegetación del este de Siberia y Alaska durante la glaciación de Wisconsin se han deducido de datos de isótopos de oxígeno de alta resolución y estratigrafía del polen . [14] [22] [23] Antes del Último Máximo Glacial, los climas en el este de Siberia fluctuaban entre condiciones que se aproximan a las actuales y períodos más fríos. Los ciclos cálidos anteriores a la LGM en el Ártico de Siberia vieron florecer las megafaunas. [14] El registro de isótopos de oxígeno del casquete glaciar de Groenlandia sugiere que estos ciclos después de unos 45.000 años AP duraron entre cientos y entre uno y dos mil años, con una mayor duración de los períodos fríos comenzando alrededor de 32.000 años cal AP. [14]El registro de polen del lago Elikchan, al norte del mar de Okhotsk , muestra un cambio marcado del polen de árboles y arbustos a polen de hierbas antes de 26k 14 C años AP, ya que la tundra de hierbas reemplazó el bosque boreal y la estepa arbustiva que ingresa al LGM. [14] Se recuperó un registro similar de polen de árboles / arbustos con polen de hierbas a medida que se acercaba el LGM cerca del río Kolyma en la Siberia ártica. [23] Se ha propuesto el abandono de las regiones del norte de Siberia debido al rápido enfriamiento o la retirada de las especies de caza con el inicio de la LGM para explicar la falta de arqueositios en esa región que datan de la LGM. [23] [24]El registro de polen del lado de Alaska muestra cambios entre hierbas / arbustos y tundra de arbustos antes de la LGM, lo que sugiere episodios de calentamiento menos dramáticos que los que permitieron la colonización de bosques en el lado siberiano. Diversas, aunque no necesariamente abundantes, megafaunas estaban presentes en esos entornos. La tundra de hierbas dominó durante la LGM, debido a las condiciones frías y secas. [22]
Los entornos costeros durante el Último Máximo Glacial fueron complejos. El descenso del nivel del mar y una protuberancia isostática equilibrada con la depresión debajo de la capa de hielo cordillerana expusieron la plataforma continental para formar una llanura costera. [25] Si bien gran parte de la llanura costera estaba cubierta por glaciares de piedemonte, se han identificado refugios no glaciares que albergan mamíferos terrestres en Haida Gwaii , la isla del Príncipe de Gales y las islas exteriores del archipiélago Alexander . [22] La llanura costera ahora sumergida tiene potencial para más refugios. [22]Los datos de polen indican principalmente vegetación de tundra herbácea / arbustiva en áreas no glaciares, con algo de bosque boreal hacia el extremo sur del rango de hielo cordillerano. [22] El entorno marino costero siguió siendo productivo, como lo indican los fósiles de pinnípedos . [25] Los bosques de algas marinas altamente productivos sobre bajíos marinos rocosos pueden haber sido un atractivo para la migración costera. [26] [27] La reconstrucción de la costa sur de Beringia también sugiere un potencial para un ambiente marino costero altamente productivo. [27]
Cambios ambientales durante la desglaciación
Los datos de polen indican un período cálido que culmina entre 14k y 11k 14 C años BP (17k-13k cal años BP) seguido de enfriamiento entre 11k-10k 14 C años BP (13k-11,5k cal años BP). [25] Las áreas costeras se desglaciaron rápidamente a medida que los glaciares alpinos costeros y luego los lóbulos del hielo cordillerano se retiraron. La retirada se aceleró a medida que aumentaba el nivel del mar y flotaba en los extremos glaciares. Se ha estimado que la cordillera de la costa estuvo completamente libre de hielo entre 16k [25] y 15k [17] años cal AP. Los organismos marinos del litoral colonizaron las costas a medida que el agua del océano reemplazó al agua de deshielo de los glaciares. El reemplazo de la tundra de hierbas / arbustos por bosques de coníferas estaba en marcha a las 12.4k 14C años BP (15k años cal BP) al norte de Haida Gwaii. El aumento eustático del nivel del mar provocó inundaciones, que se aceleraron a medida que aumentaba la velocidad. [25]
Las capas de hielo del interior de Cordilleran y Laurentide se retiraron más lentamente que los glaciares costeros. La apertura de un corredor sin hielo no ocurrió hasta después de 13k a 12k años cal AP. [15] [16] [17] El entorno inicial del corredor libre de hielo estuvo dominado por el deshielo glacial y el agua de deshielo, con lagos con represas de hielo e inundaciones periódicas por la liberación de agua de deshielo. [15] La productividad biológica del paisaje desglaciado se ganó lentamente. [17] La viabilidad más temprana posible del corredor libre de hielo como ruta de migración humana se ha estimado en 11,5 mil años cal AP. [17]
Los bosques de abedules avanzaban a través de la antigua tundra de hierbas en Beringia hacia 14.3ka 14 C años AP (17k años cal AP) en respuesta a la mejora climática, lo que indica una mayor productividad del paisaje. [23]
Los análisis de biomarcadores y microfósiles conservados en sedimentos del lago E5 y del lago Burial en el norte de Alaska sugieren que los primeros humanos quemaron paisajes de Beringia hace 34.000 años. [28] [29] Los autores de estos estudios sugieren que el fuego se utilizó como medio para cazar megafauna.
Cronología, motivos y fuentes de la migración

La comunidad arqueológica está en general de acuerdo en que los antepasados de los pueblos indígenas de las Américas de registro histórico ingresaron a las Américas al final del Último Máximo Glacial (LGM), poco después de hace 20.000 años, con presencia arqueológica comprobada poco después de hace 16.000 años. .
Sigue habiendo incertidumbres con respecto a la datación precisa de sitios individuales y con respecto a las conclusiones extraídas de los estudios de genética de poblaciones de los nativos americanos contemporáneos. También es una pregunta abierta si esta migración post-LGM representó el primer poblamiento de las Américas, o si hubo una migración anterior, pre-LGM que llegó a Sudamérica hace 40.000 años.
Cronología
A principios del siglo XXI, los modelos de cronología de la migración se dividen en dos enfoques generales. [30] [31]
La primera es la teoría de la cronología corta , según la cual la primera migración ocurrió después del Último Máximo Glacial, que entró en declive después de hace unos 19.000 años, [20] y luego fue seguida por sucesivas oleadas de inmigrantes. [32]
La segunda teoría es la teoría de la cronología larga, que propone que el primer grupo de personas ingresó a las Américas en una fecha mucho más temprana, posiblemente antes de hace 40.000 años, [33] [34] [35] seguido de una segunda ola de inmigrantes mucho más tardía. . [31] [36]
La teoría de Clovis First , que dominó el pensamiento sobre la antropología del Nuevo Mundo durante gran parte del siglo XX, fue cuestionada por la datación segura de los arqueositos en las Américas hasta hace 13.000 años en la década de 2000. [15] [16] [17] [37] [38] El escenario de "cronología corta", a la luz de esto, se refiere a un poblamiento de las Américas poco después de hace 19.000 años, mientras que el escenario de "cronología larga" permite pre -Presencia LGM, hace unos 40.000 años.
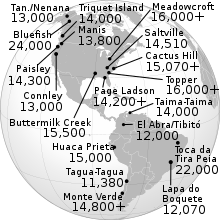
Los arqueositios de las Américas con las fechas más antiguas que han ganado una amplia aceptación son todos compatibles con una edad de aproximadamente 15.000 años. Esto incluye el complejo Buttermilk Creek en Texas, [39] el sitio Meadowcroft Rockshelter en Pensilvania y el sitio Monte Verde en el sur de Chile. [38] La evidencia arqueológica de la gente anterior a Clovis apunta a que el sitio Topper de Carolina del Sur tiene 16.000 años de antigüedad, en un momento en que el máximo glacial habría permitido teóricamente costas más bajas.
A menudo se ha sugerido que un corredor sin hielo, en lo que ahora es el oeste de Canadá , habría permitido la migración antes del comienzo del Holoceno , pero un estudio de 2016 ha argumentado en contra de esto, sugiriendo que el poblamiento de América del Norte a través de dicho corredor Es poco probable que sea significativamente anterior a los primeros sitios de Clovis. El estudio concluye que el corredor libre de hielo en lo que ahora es Alberta y Columbia Británica "fue tomado gradualmente por un bosque boreal dominado por abetos y pinos" y que "la gente de Clovis probablemente vino del sur, no del norte, tal vez siguiendo a animales salvajes como el bisonte ". [40] [41]Una hipótesis alternativa para el poblamiento de América es la migración costera , que puede haber sido factible a lo largo de la costa desglaciada (pero ahora sumergida) del noroeste del Pacífico desde hace unos 16.000 años.
Evidencia de la presencia humana pre-LGM
!["Mapas que representan cada fase de las primeras migraciones humanas de tres pasos para el poblamiento de las Américas. (A) Expansión gradual de la población de los antepasados amerindios de su acervo genético de Asia Central y Oriental (flecha azul). (B) Ocupación protoamerindia de Beringia con poco o ningún crecimiento de población durante ~ 20.000 años. (C) Rápida colonización del Nuevo Mundo por un grupo fundador que migró hacia el sur a través del corredor interior libre de hielo entre las capas de hielo del este de Laurentide y el oeste de la Cordillera (flecha verde) y / o a lo largo de la costa del Pacífico (flecha roja). En (B), el fondo marino expuesto se muestra en su mayor extensión durante el último máximo glacial en hace 20 a 18 000 años [25]. En (A) y (C), el El fondo marino expuesto se representa hace 40.000 años y hace 16.000 años, cuando los niveles del mar prehistóricos eran comparables.En la esquina inferior izquierda se presenta una versión reducida de Beringia (reducción del 60% de A – C). Este mapa más pequeño destaca el estrecho de Bering que ha separado geográficamente el Nuevo Mundo de Asia desde hace ≈11–10.000 años ".](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d7/Journal.pone.0001596.g004.png/280px-Journal.pone.0001596.g004.png)
Se ha propuesto la migración anterior al último máximo glacial a través de Beringia hacia las Américas para explicar las supuestas edades pre-LGM de las arqueositas en las Américas, como Bluefish Caves [34] y Old Crow Flats [35] en el territorio de Yukon , y Meadowcroft Rock Shelter en Pensilvania. [31] [36]
En Old Crow Flats, se han encontrado huesos de mamut que están rotos de formas distintivas que indican una carnicería humana. Las fechas de radiocarbono en estos varían entre 25.000 y 40.000 años AP. Además, se han encontrado microescamas de piedra en el área que indican la producción de herramientas. [43]
Anteriormente, las interpretaciones de las marcas de carnicero y la asociación geológica de huesos en los sitios Bluefish Cave y Old Crow Flats, y el sitio Bonnet Plume relacionado, se han puesto en duda. [44]
Además de los sitios arqueológicos en disputa, se ha encontrado apoyo para la presencia humana pre-LGM en los registros de sedimentos de lagos del norte de Alaska. Los análisis de biomarcadores y microfósiles de sedimentos del lago E5 y del lago Burial sugieren la presencia humana en el este de Beringia hace 34.000 años. [28] [29] Estos análisis son de hecho convincentes en el sentido de que corroboran las inferencias hechas a partir de los sitios Bluefish Cave y Old Crow Flats.
En 2020, surgieron pruebas de un nuevo sitio pre-LGM en el centro-norte de México . La cueva de Chiquihuite , un sitio arqueológico en el estado de Zacatecas , data de hace 26,000 años AP basado en numerosos artefactos líticos descubiertos allí. [45]
La presencia humana pre-LGM en América del Sur se basa en parte en la cronología del controvertido refugio rocoso Pedra Furada en Piauí , Brasil . Un estudio de 2003 fechó la evidencia del uso controlado del fuego antes de hace 40.000 años. [46] Se han aportado pruebas adicionales a partir de la morfología del fósil Mujer Luzia , que se describió como australoide . Esta interpretación fue cuestionada en una revisión de 2003 que concluyó que las características en cuestión también podrían haber surgido por deriva genética. [47] En noviembre de 2018, científicos de la Universidad de São Paulo y la Universidad de Harvardpublicó un estudio que contradice el supuesto origen australo-melanesio de Luzia. Usando la secuenciación de ADN, los resultados mostraron que Luzia era completamente amerindia, genéticamente. [48] [49]
Las edades de los primeros artefactos identificados positivamente en el sitio de Meadowcroft se encuentran de manera segura dentro del período posterior a la LGM (13.8k – 18.5k años cal BP ). [37] [50]
En el sur de California , en el sitio Cerutti Mastodon , se han encontrado piedras descritas como probables herramientas, martillos y yunques , que están asociadas con un esqueleto de mastodonte que parecía haber sido procesado por humanos. El esqueleto del mastodonte se fechó mediante análisis radiométrico de torio-230 / uranio, utilizando modelos de datación por difusión-adsorción-desintegración, hasta hace 130,7 ± 9,4 mil años. [51] No se encontraron huesos humanos y la reacción de los expertos fue mixta; Las afirmaciones sobre herramientas y procesamiento de huesos fueron calificadas de "no plausibles" por el Prof. Tom Dillehay . [52]
El sitio del cuerno de rinoceronte del río Yana (RHS) ha fechado la ocupación humana del este de Siberia ártica en 27k 14 C años AP (31,3k años cal AP). [53] Esa fecha ha sido interpretada por algunos como evidencia de que la migración a Beringia era inminente, dando crédito a la ocupación de Beringia durante la LGM. [54] [55] Sin embargo, la fecha de Yana RHS es desde el comienzo del período de enfriamiento que condujo al LGM. [14] Sin embargo, una recopilación de fechas de arqueositas en todo el este de Siberia sugiere que el período de enfriamiento provocó una retirada de los humanos hacia el sur. [23] [24]La evidencia lítica anterior a la LGM en Siberia indica un estilo de vida asentado que se basó en los recursos locales, mientras que la evidencia lítica posterior a la LGM indica un estilo de vida más migratorio. [24]
La arqueosita más antigua en el lado de Alaska de Beringia data de 12k 14 C años AP (14k años cal AP). [23] [56] Es posible que una pequeña población fundadora hubiera entrado en Beringia antes de esa época. Sin embargo, faltan arqueositas que datan más cerca del Último Máximo Glacial en el lado siberiano o en el lado de Alaska de Beringia. Los análisis de biomarcadores y microfósiles de sedimentos del lago E5 y del lago Burial en el norte de Alaska sugieren la presencia humana en el este de Beringia hace 34.000 años. [28] Se ha sugerido que estos análisis sedimentarios son los únicos restos posiblemente recuperables de humanos que vivieron en Alaska durante el último período glacial. [29]
Estimaciones de la edad genómica
Los estudios de genética amerindia han utilizado técnicas analíticas de alta resolución aplicadas a muestras de ADN de poblaciones nativas americanas modernas y asiáticas consideradas como sus poblaciones de origen para reconstruir el desarrollo de haplogrupos de ADN del cromosoma Y humano ( haplogrupos de yADN ) y haplogrupos de ADN mitocondrial humano (haplogrupos de ADNmt). característica de las poblaciones nativas americanas. [33] [54] [55]Se utilizaron modelos de tasas de evolución molecular para estimar las edades en las que los linajes de ADN de los nativos americanos se ramificaron de sus linajes parentales en Asia y para deducir las edades de los eventos demográficos. Un modelo (Tammetal 2007) basado en los haplotipos de ADNmt de los nativos americanos (Figura 2) propone que la migración a Beringia ocurrió entre 30k y 25k años cal AP, y que la migración a las Américas se produjo alrededor de 10k a 15k años después del aislamiento de la pequeña población fundadora . [54] Otro modelo (Kitchen et al. 2008) propone que la migración a Beringia ocurrió aproximadamente 36.000 años cal AP, seguida de 20.000 años de aislamiento en Beringia. [55]Un tercer modelo (Nomatto et al. 2009) propone que la migración a Beringia ocurrió entre 40k y 30k años cal AP, con una migración anterior a LGM en las Américas seguida por el aislamiento de la población del norte luego del cierre del corredor libre de hielo. [33] Skoglund y Reich (2016) encontraron evidencia de la mezcla australo-melanesia en poblaciones amazónicas. [57]
Un estudio de la diversificación de ADNmt haplogrupos C y D del sur de Siberia y Asia oriental, respectivamente, sugiere que el linaje de los padres (subhaplogroup D4h) de subhaplogroup D4h3, un linaje encontrado entre los nativos americanos y los chinos Han, [58] [59] emergió alrededor de 20.000 años cal. BP, lo que limita la aparición de D4h3 a post-LGM. [60] Las estimaciones de edad basadas en la diversidad de microsatélites del cromosoma Y sitúan el origen del Haplogrupo estadounidense Q1a3a (ADN-Y) entre 10.000 y 15.000 años cal BP. [61] Se puede obtener una mayor coherencia de los modelos de tasa de evolución molecular del ADN entre sí y con los datos arqueológicos mediante el uso de ADN fósil fechado para calibrar las tasas de evolución molecular. [58]
Poblaciones de origen
Existe un acuerdo general entre los antropólogos de que las poblaciones de origen de la migración a las Américas se originaron en un área en algún lugar al este del río Yenisei ( Lejano Oriente ruso ). La ocurrencia común del ADNmt haplogrupos A, B, C, y D en las poblaciones asiáticas y nativos americanos del este tiempo se ha reconocido, junto con la presencia del haplogrupo X . [62] En conjunto, la mayor frecuencia de los cuatro haplogrupos asociados de nativos americanos se produce en la región de Altai - Baikal en el sur de Siberia. [63] Algunos subcladesde C y D más cercanos a los subclades de los nativos americanos se encuentran entre las poblaciones de Mongolia, Amur, Japón, Corea y Ainu. [62] [64]
Un estudio de 2019 sugirió que los nativos americanos son los parientes vivos más cercanos a los fósiles de 10.000 años encontrados cerca del río Kolyma en el noreste de Siberia. [sesenta y cinco]
Modelos genómicos humanos
The development of high-resolution genomic analysis has provided opportunities to further define Native American subclades and narrow the range of Asian subclades that may be parent or sister subclades. For example, the broad geographic range of haplogroup X has been interpreted as allowing the possibility of a western Eurasian, or even a European source population for Native Americans, as in the Solutrean hypothesis, or suggesting a pre-Last Glacial Maximum migration into the Americas.[62] The analysis of an ancient variant of haplogroup X among aboriginals of the Altai region indicates common ancestry with the European strain rather than descent from the European strain.[63]Una mayor división de los subclades X ha permitido la identificación del subhaplogrupo X2a, que se considera específico de los nativos americanos. [54] [59] Con una mayor definición de subclades relacionados con las poblaciones de nativos americanos, los requisitos para el muestreo de poblaciones asiáticas para encontrar los subclades más estrechamente relacionados se vuelven más específicos. Los subhaplogrupos D1 y D4h3 se han considerado específicos de los nativos americanos en función de su ausencia entre una gran muestra de poblaciones consideradas como descendientes potenciales de las poblaciones de origen, en una amplia zona de Asia. [54] Entre las 3764 muestras, la región de Sakhalin - bajo Amur estuvo representada por 61 Oroks . [54]En otro estudio, se identificó el subhaplogrupo D1a entre los Ulchis de la región del bajo río Amur (4 entre 87 muestreados, o 4,6%), junto con el subhaplogrupo C1a (1 entre 87, o 1,1%). [64] El subhaplogrupo C1a se considera un clado hermano cercano del subhaplogrupo C1b de los nativos americanos. [64]
El subhaplogrupo D1a también se ha encontrado entre los esqueletos Jōmon antiguos de Hokkaido [66]. Los Ainu modernos son considerados descendientes de los Jōmon. [66] La ocurrencia de los subhaplogrupos D1a y C1a en la región del bajo Amur sugiere una población fuente de esa región distinta de las poblaciones fuente de Altai-Baikal, donde el muestreo no reveló esos dos subclades particulares. [64] Las conclusiones con respecto al subhaplogrupo D1 que indican poblaciones de origen potenciales en las áreas inferiores de Amur [64] y Hokkaido [66] contrastan con el modelo de migración de origen único. [33] [54] [55]
Se ha identificado el subhaplogrupo D4h3 entre los chinos han . [58] [59] El subhaplogrupo D4h3 de China no tiene la misma implicación geográfica que el subhaplotipo D1a de Amur-Hokkaido, por lo que sus implicaciones para los modelos fuente son más especulativas. Se cree que su linaje parental, el subhaplotipo D4h, surgió en el este de Asia, en lugar de Siberia, alrededor de 20.000 años cal. AP. [60] El subhaplogrupo D4h2, un clado hermano de D4h3, también se ha encontrado entre los esqueletos de Jōmon de Hokkaido. [67] D4h3 tiene un rastro costero en las Américas. [59]
El contraste entre los perfiles genéticos de los esqueletos de Hokkaido Jōmon y los Ainu modernos ilustra otra incertidumbre en los modelos fuente derivados de muestras de ADN modernas: [66]
Sin embargo, probablemente debido al pequeño tamaño de la muestra o la estrecha consanguinidad entre los miembros del sitio, las frecuencias de los haplogrupos en los esqueletos de Funadomari eran bastante diferentes de las de cualquier población moderna, incluido Hokkaido Ainu, que ha sido considerado descendiente directo de Hokkaido. Gente jomon.
Los descendientes de las poblaciones de origen con la relación más cercana al perfil genético desde el momento en que ocurrió la diferenciación no son obvios. Se puede esperar que los modelos de población de origen se vuelvan más sólidos a medida que se compilen más resultados, se comprenda mejor la herencia de los candidatos proxy modernos y se encuentre y considere el ADN fósil en las regiones de interés.
Genómica HTLV-1
El virus linfotrófico de células T humanas 1 ( HTLV-1 ) es un virus que se transmite a través del intercambio de fluidos corporales y de madre a hijo a través de la leche materna. La transmisión de madre a hijo imita un rasgo hereditario, aunque dicha transmisión de los portadores maternos es inferior al 100%. [68] Se ha cartografiado el genoma del virus HTLV, lo que permite la identificación de cuatro cepas principales y el análisis de su antigüedad a través de mutaciones. Las concentraciones geográficas más altas de la cepa HLTV-1 se encuentran en África subsahariana y Japón. [69] En Japón, ocurre en su mayor concentración en Kyushu . [69] También está presente entre los descendientes de africanos y las poblaciones nativas en la región del Caribe y América del Sur. [69]Es raro en América Central y América del Norte. [69] Se ha considerado que su distribución en las Américas se debe a la importación con el comercio de esclavos. [70]
Los Ainu han desarrollado anticuerpos contra HTLV-1, lo que indica su endemicidad para los Ainu y su antigüedad en Japón. [71] Se ha definido e identificado un subtipo "A" entre los japoneses (incluidos los ainu) y entre los aislados caribeños y sudamericanos. [72] Se ha identificado un subtipo "B" en Japón y la India. [72] En 1995, se descubrió que los nativos americanos de la costa de Columbia Británica tenían ambos subtipos A y B. [73] Se informó que los especímenes de médula ósea de una momia andina de unos 1500 años de antigüedad mostraron la presencia del subtipo A. [74] El hallazgo generó controversia, con el argumento de que la muestra de ADN no estaba lo suficientemente completa para la conclusión y que el resultado reflejaba una contaminación moderna.[75] Sin embargo, un nuevo análisis indicó que las secuencias de ADN eran consistentes con, pero no definitivamente del "clado cosmopolita" (subtipo A). [75] La presencia de subtipos A y B en las Américas sugiere una población de origen nativo americano relacionada con los antepasados Ainu, los Jōmon.
Antropología física
Esqueletos paleoamericanos en las Américas como el Hombre Kennewick (Estado de Washington), el esqueleto Hoya Negro (Yucatán), la Mujer Luzia y otros cráneos del sitio Lagoa Santa (Brasil), la Mujer Buhl (Idaho), la Mujer Peñón III , [76] dos cráneos del sitio de Tlapacoya (Ciudad de México), [76] y 33 cráneos de Baja California [77] han exhibido rasgos craneofaciales distintos de la mayoría de los nativos americanos modernos, lo que llevó a los antropólogos físicos a la opinión de que algunos paleoamericanos eran de origen australoide más que siberiano. [78]El rasgo distintivo medido más básico es la dolicocefalia del cráneo. Algunas cepas modernas como los pericúes de Baja California y los fueguinos de Tierra del Fuego que exhiben mismo rasgo morfológico. [77] Otros antropólogos defienden una hipótesis alternativa de que la evolución de un fenotipo original de Beringio dio lugar a una morfología distinta que era similar en todos los cráneos paleoamericanos conocidos, seguida de una convergencia posterior hacia el fenotipo nativo americano moderno. [79] [80] Los estudios genéticos apoyan un origen australoide. [57]
Un informe publicado en el American Journal of Physical Anthropologyen enero de 2015 revisó la variación craneofacial centrándose en las diferencias entre los nativos americanos tempranos y tardíos y las explicaciones de estas basadas en la morfología del cráneo o la genética molecular. Los argumentos basados en la genética molecular han aceptado en su mayoría, según los autores, una única migración desde Asia con una probable pausa en Berengia, más un flujo genético bidireccional posterior. Los estudios que se centran en la morfología craneofacial han argumentado que los restos paleoamericanos "han sido descritos como mucho más cercanos a las poblaciones africanas y australo-melanesias que a la serie moderna de nativos americanos", lo que sugiere dos entradas en las Américas, una de las cuales ocurrió antes de un distintivo de Asia oriental. morfología desarrollada (referido en el artículo como el "Modelo de dos componentes". Un tercer modelo, el "Flujo genético recurrente"El modelo [RGF], intenta reconciliar los dos, argumentando que el flujo de genes circumarcticos después de la migración inicial podría explicar los cambios morfológicos. Reevalúa específicamente el informe original sobre el esqueleto de Hoya Negro que apoyaba el modelo RGF, los autores no estaban de acuerdo con la conclusión original que sugería que la forma del cráneo no coincidía con la de los nativos americanos modernos, argumentando que el "cráneo cae en una subregión del morfoespacio ocupado tanto por los paleoamericanos como por algunos nativos americanos modernos ".los autores no estuvieron de acuerdo con la conclusión original que sugería que la forma del cráneo no coincidía con la de los nativos americanos modernos, argumentando que "el cráneo cae en una subregión del morfoespacio ocupado tanto por los paleoamericanos como por algunos nativos americanos modernos".los autores no estuvieron de acuerdo con la conclusión original que sugería que la forma del cráneo no coincidía con la de los nativos americanos modernos, argumentando que "el cráneo cae en una subregión del morfoespacio ocupado tanto por los paleoamericanos como por algunos nativos americanos modernos".[81]
Puntas con tallo
Las puntas madre son una tecnología lítica distinta de los tipos Beringian y Clovis. Tienen una distribución que va desde la costa este de Asia hasta la costa del Pacífico de América del Sur. [26] La aparición de puntas derivadas se remonta a Corea durante el Paleolítico superior. [82] El origen y la distribución de las puntas derivadas se han interpretado como un marcador cultural relacionado con una población de origen de la costa este de Asia. [26]
Rutas de migración
Ruta interior

Históricamente, las teorías sobre la migración a las Américas han girado en torno a la migración desde Beringia hacia el interior de América del Norte. El descubrimiento de artefactos en asociación con restos de fauna del Pleistoceno cerca de Clovis, Nuevo México, a principios de la década de 1930, requirió la extensión del período de tiempo para el asentamiento de América del Norte al período durante el cual los glaciares aún eran extensos. Eso llevó a la hipótesis de una ruta de migración entre las capas de hielo Laurentide y Cordilleran para explicar el asentamiento temprano. El sitio de Clovis fue sede de una tecnología lítica caracterizada por puntas de lanza con una muesca, o flauta, donde la punta estaba unida al eje. Un complejo lítico caracterizado por la Punta ClovisPosteriormente se identificó tecnología en gran parte de América del Norte y América del Sur. La asociación de la tecnología del complejo Clovis con restos de fauna del Pleistoceno tardío llevó a la teoría de que marcó la llegada de los cazadores de caza mayor que emigraron de Beringia y luego se dispersaron por las Américas, también conocida como la teoría de Clovis First.
La datación por radiocarbono reciente de los sitios de Clovis ha arrojado edades de 11.1k a 10.7k 14 C años AP (13k a 12.6k años cal AP), algo más tarde que las fechas derivadas de técnicas más antiguas. [83] La reevaluación de las fechas de radiocarbono anteriores llevó a la conclusión de que no menos de 11 de los 22 sitios Clovis con fechas de radiocarbono son "problemáticos" y deben ignorarse, incluido el sitio tipo en Clovis, Nuevo México. La datación numérica de los sitios de Clovis ha permitido comparar las fechas de Clovis con las fechas de otros arqueositios de América y de la apertura del corredor libre de hielo. Ambos conducen a desafíos importantes para la teoría de Clovis First. El sitio de Monte Verde en el sur de Chile se ha fechado en 14.8k años cal AP. [38]El sitio de la cueva de Paisley en el este de Oregon arrojó una fecha de 14 C de 12.4k años (14.5k años cal) BP, en un coprolito con ADN humano y fechas 14 C de 11.3k-11k (13.2k-12.9k años cal) BP en horizontes que contienen puntas derivadas del oeste. [84] Los horizontes de artefactos con conjuntos líticos no clovis y edades anteriores a Clovis ocurren en el este de América del Norte, aunque las edades máximas tienden a estar poco restringidas. [37] [50]
Los hallazgos geológicos sobre el momento del corredor libre de hielo también desafían la noción de que Clovis y la ocupación humana de las Américas antes de Clovis fue el resultado de la migración a través de esa ruta siguiendo el Último Máximo Glacial . El cierre del corredor antes del LGM puede acercarse a los 30.000 años cal BP y las estimaciones de la retirada del hielo del corredor están en el rango de 12 a 13.000 años cal BP. [15] [16] [17] La viabilidad del corredor como ruta de migración humana se ha estimado en 11.5k años cal AP, más tarde que las edades de los sitios Clovis y pre-Clovis. [17] Las arqueositas datadas de Clovis sugieren una expansión de la cultura Clovis de sur a norte. [15]
Se ha propuesto la migración previa al último máximo glacial hacia el interior para explicar las edades anteriores a Clovis de los arqueositos en las Américas, [31] [36] aunque los sitios anteriores a Clovis como Meadowcroft Rock Shelter, [37] [50] Monte Verde, [38] y Paisley Cave no han arrojado edades pre-LGM confirmadas.
Propuesta de familia lingüística dené-yeniseiana
Una relación entre las lenguas na-dené de América del Norte (como el navajo y el apache) y las lenguas yeniseianas de Siberia fue propuesta por primera vez en 1923 y desarrollada por otros. Edward Vajda realizó un estudio detallado que se publicó en 2010. [85] Esta teoría recibió el apoyo de muchos lingüistas, y los estudios arqueológicos y genéticos le proporcionaron más apoyo.
La tradición de las herramientas pequeñas del Ártico de Alaska y el Ártico canadiense puede haberse originado en el este de Siberia hace unos 5.000 años. Esto está relacionado con los antiguos pueblos paleo-esquimales del Ártico, la cultura que se desarrolló hacia el 2500 a. C.
La fuente de la tradición de la Herramienta Pequeña del Ártico puede haber sido la secuencia de la cultura Syalakh- Bel'kachi- Ymyakhtakh de Siberia Oriental, que data de 6.500 - 2.800 calBP. [86]
La ruta interior es consistente con la expansión del grupo lingüístico Na-Dene [85] y el subhaplogrupo X2a en las Américas después de la primera migración paleoamericana. [59]
Sin embargo, algunos estudiosos sugieren que los antepasados de los norteamericanos occidentales que hablaban lenguas na-denés hicieron una migración costera en barco. [87]
Ruta costera del Pacífico
La teoría de la migración costera del Pacífico propone que las personas llegaron primero a las Américas viajando por agua, siguiendo las costas desde el noreste de Asia hasta las Américas, propuesto originalmente en 1979 por Knute Fladmark como una alternativa a la migración hipotética a través de un corredor interior sin hielo. [88] Este modelo ayudaría a explicar la rápida propagación a sitios costeros extremadamente distantes de la región del Estrecho de Bering, incluidos sitios como Monte Verde en el sur de Chile y Taima-Taima en el oeste de Venezuela .
La hipótesis de la migración marina muy similar es una variante de la migración costera; esencialmente, su única diferencia es que postula que los barcos eran el principal medio de viaje. El uso propuesto de botes agrega una medida de flexibilidad a la cronología de la migración costera, porque entonces no se requeriría una costa continua sin hielo (16-15,000 años calibrados AP): los migrantes en botes podrían haber pasado fácilmente por encima de las barreras de hielo y asentarse en refugios costeros dispersos, antes de que se completara la desglaciación de la ruta terrestre costera. Una población de origen con competencia marítima en la costa este de Asia es una parte esencial de la hipótesis de la migración marina. [26] [27]
Un artículo de 2007 en el Journal of Island and Coastal Archaeology propuso una "hipótesis de la carretera de algas marinas", una variante de la migración costera basada en la explotación de bosques de algas marinas a lo largo de gran parte del borde del Pacífico desde Japón hasta Beringia, el noroeste del Pacífico y California, y hasta la Costa Andina de América del Sur. Una vez que las costas de Alaska y Columbia Británica se desglaciaron hace unos 16.000 años, estos hábitats de bosques de algas marinas (junto con estuarios, manglares y arrecifes de coral) habrían proporcionado un corredor de migración ecológicamente homogéneo, completamente al nivel del mar y esencialmente sin obstrucciones. Un análisis de ADN de plantas y animales de 2016 sugiere que una ruta costera era factible. [89] [90]
El subhaplogrupo mitocondrial D4h3a , un subclade raro de D4h3 que se encuentra a lo largo de la costa oeste de las Américas, ha sido identificado como un clado asociado con la migración costera. [59] Este haplogrupo se encontró en un esqueleto conocido como Anzick-1 , encontrado en Montana en estrecha asociación con varios artefactos de Clovis, fechados hace 12.500 años. [91]
Problemas con la evaluación de modelos de migración costera
Los modelos de migración costera ofrecen una perspectiva diferente sobre la migración al Nuevo Mundo, pero no están exentos de problemas: uno de esos problemas es que los niveles globales del mar han aumentado más de 120 metros (390 pies) [92] desde el final del último período glacial, y esto ha sumergido las antiguas costas que la gente marítima habría seguido en las Américas. Encontrar sitios asociados con migraciones costeras tempranas es extremadamente difícil, y la excavación sistemática de cualquier sitio que se encuentre en aguas más profundas es desafiante y costoso. Las estrategias para encontrar los sitios de migración más tempranos incluyen la identificación de sitios potenciales en paleoshorelines sumergidos, buscando sitios en áreas elevadas por tectónica o rebote isostático, y buscando sitios ribereños en áreas que pueden haber atraído a migrantes costeros. [26] [93] Por otro lado, existe evidencia de tecnologías marinas encontradas en las colinas de las Islas del Canal de California , alrededor del año 10.000 a. C. [94] Si hubo una migración costera temprana antes de Clovis, siempre existe la posibilidad de una "colonización fallida". Otro problema que surge es la falta de evidencia sólida encontrada para una teoría de "cronología larga". Ningún sitio ha producido aún una cronología coherente con más de 12.500 años de radiocarbono (~ 14.500 años calendario [ cita requerida ]).), pero la investigación en América del Sur relacionada con la posibilidad de migraciones costeras tempranas ha sido limitada.
Ver también
- Migraciones humanas tempranas
- Lista de primeros asentamientos humanos
- Historia genética de los pueblos indígenas de las Américas
- Teorías de contacto transoceánicas precolombinas
Referencias
- ^ Pringle, Heather (8 de marzo de 2017). "¿Qué sucede cuando un arqueólogo desafía el pensamiento científico convencional?" . Smithsonian .
- ^ Fagan, Brian M. y Durrani, Nadia (2016). Prehistoria mundial: una breve introducción . Routledge. pag. 124. ISBN 978-1-317-34244-1.
- ^ a b Goebel, Ted; Waters, Michael R .; O'Rourke, Dennis H. (2008). "La dispersión del Pleistoceno tardío de los humanos modernos en las Américas" (PDF) . Ciencia . 319 (5869): 1497–1502. Código bibliográfico : 2008Sci ... 319.1497G . CiteSeerX 10.1.1.398.9315 . doi : 10.1126 / science.1153569 . PMID 18339930 . S2CID 36149744 . Archivado desde el original el 2 de enero de 2014 . Consultado el 5 de febrero de 2010 . CS1 maint: bot: original URL status unknown (link)
- ^ Zimmer, Carl (3 de enero de 2018). "En los huesos de un niño enterrado, signos de una masiva migración humana a las Américas" . The New York Times . Consultado el 3 de enero de 2018 .
- ^ Moreno-Mayar, JV; Potter, BA; Vinner, L; et al. (2018). "El genoma de Alaska del Pleistoceno terminal revela la primera población fundadora de nativos americanos" (PDF) . Naturaleza . 553 (7687): 203–207. Código Bib : 2018Natur.553..203M . doi : 10.1038 / nature25173 . PMID 29323294 . S2CID 4454580 .
- ^ Waguespack, Nicole (2012). "Paleoindios tempranos, de la colonización a Folsom" . En Timothy R. Pauketat (ed.). El Manual de Oxford de Arqueología de América del Norte . Prensa de la Universidad de Oxford. págs. 86–95. ISBN 978-0-19-538011-8.
- ^ Kornfeld, Marcel y Politis, Gustavo G. (2014). "En las Américas: los primeros cazadores-recolectores en un continente vacío" . En Vicki Cummings; Peter Jordan y Marek Zvelebil (eds.). El Manual de Oxford de Arqueología y Antropología de Cazadores-Recolectores . Prensa de la Universidad de Oxford. pag. 406. ISBN 978-0-19-102526-6.
- ^ Ash, Patricia J. y Robinson, David J. (2011). El surgimiento de los humanos: una exploración de la línea de tiempo evolutiva . John Wiley e hijos . pag. 289. ISBN 978-1-119-96424-7.
- ^ Roberts, Alice (2010). El increíble viaje humano . A&C Negro. págs. 101-103. ISBN 978-1-4088-1091-0.
- ^ Ardelean, Ciprian F .; Becerra-Valdivia, Lorena; Pedersen, Mikkel Winther; Schwenninger, Jean-Luc; Oviatt, Charles G .; Macías-Quintero, Juan I .; Arroyo-Cabrales, Joaquín; Sikora, Martin; Ocampo-Díaz, Yam Zul E .; Rubio-Cisneros, Igor I .; Watling, Jennifer G .; De Medeiros, Vanda B .; De Oliveira, Paulo E .; Barba-Pingarón, Luis; Ortiz-Butrón, Agustín; Blancas-Vázquez, Jorge; Rivera-González, Irán; Solís-Rosales, Corina; Rodríguez-Ceja, María; Gandy, Devlin A .; Navarro-Gutiérrez, Zamara; de la Rosa-Díaz, Jesús J .; Huerta-Arellano, Vladimir; Marroquín-Fernández, Marco B .; Martínez-Riojas, L. Martín; López-Jiménez, Alejandro; Higham, Thomas; Willerslev, Eske (2020). "Evidencia de ocupación humana en México en torno al Último Máximo Glacial" . Naturaleza . 584(7819): 87–92. Código Bib : 2020Natur.584 ... 87A . doi : 10.1038 / s41586-020-2509-0 . PMID 32699412 . S2CID 220697089 .
- ↑ Becerra-Valdivia, Lorena; Higham, Thomas (2020). "El momento y el efecto de las primeras llegadas humanas a América del Norte". Naturaleza . 584 (7819): 93–97. Código Bibliográfico : 2020Natur.584 ... 93B . doi : 10.1038 / s41586-020-2491-6 . PMID 32699413 . S2CID 220715918 .
- ^ Gruhn, Ruth (22 de julio de 2020). “Crece la evidencia de que el poblamiento de las Américas comenzó hace más de 20.000 años” . Naturaleza . 584 (7819): 47–48. Código Bib : 2020Natur.584 ... 47G . doi : 10.1038 / d41586-020-02137-3 . PMID 32699366 . S2CID 220717778 .
- ^ Somerville, Andrew D .; Casar, Isabel; Arroyo-Cabrales, Joaquín (2021). "Nuevas Edades de Radiocarbono AMS de los Niveles Precerámicos de la Cueva de Coxcatlán, Puebla, México: ¿Una Ocupación Pleistocena del Valle de Tehuacán?" . Antigüedad latinoamericana : 1-15. doi : 10.1017 / laq.2021.26 .
- ^ a b c d e f g h Brigham-Grette, Julie; Lozhkin, Anatoly V .; Anderson, Patricia M. y Glushkova, Olga Y. (2004). "Condiciones paleoambientales en West Beringia antes del último máximo glacial" . En DB Madsen (ed.). Entrando en América: el noreste de Asia y Beringia antes del último máximo glacial . Prensa de la Universidad de Utah. ISBN 978-0-87480-786-8.
- ↑ a b c d e f g Jackson, Lionel E., Jr. y Wilson, Michael C. (febrero de 2004). "El corredor libre de hielo revisitado" . Geotimes . Instituto Geológico Americano .
- ^ a b c d e Jackson, LE Jr .; Phillips, FM; Shimamura, K. y Little, EC (1997). "Cosmogenic 36 Cl data del tren Foothills Erratics, Alberta, Canadá". Geología . 25 (3): 195-198. Código Bibliográfico : 1997Geo .... 25..195J . doi : 10.1130 / 0091-7613 (1997) 025 <0195: ccdotf> 2.3.co; 2 .
- ^ a b c d e f g h i j k Mandryk, Carole AS; Josenhans, Heiner; Fedje, Daryl W. y Mathewes, Rolf W. (enero de 2001). "Paleoambientes del Cuaternario tardío del noroeste de América del Norte: implicaciones para las rutas de migración hacia el interior frente a las costeras" . Reseñas de ciencias cuaternarias . 20 (1): 301–314. Código Bibliográfico : 2001QSRv ... 20..301M . doi : 10.1016 / s0277-3791 (00) 00115-3 .
- ^ a b Dyke, AS; Moore, A. y Robertson, L. (2003). Desglaciación de América del Norte (Informe). Archivo abierto 1574. Servicio geológico de Canadá . doi : 10.4095 / 214399 .
- ^ a b Stand, Derek B .; Troost, Kathy Goetz; Clague, John J. y Waitt, Richard B. (2003). "La capa de hielo de la Cordillera". El período cuaternario en los Estados Unidos . Desarrollos en Ciencias Cuaternarias. 1 . págs. 17–43. doi : 10.1016 / S1571-0866 (03) 01002-9 . ISBN 978-0-4445-1470-7.
- ^ a b Blaise, B .; Clague, JJ y Mathewes, RW (1990). "Tiempo de máxima glaciación tardía de Wisconsin, costa oeste de Canadá". Investigación Cuaternaria . 34 (3): 282–295. Código Bibliográfico : 1990QuRes..34..282B . doi : 10.1016 / 0033-5894 (90) 90041-i .
- ^ Misarti, Nicole; Finney, Bruce P .; Jordan, James W .; et al. (10 de agosto de 2012). "Retiro temprano del complejo glaciar de la península de Alaska y las implicaciones para las migraciones costeras de los primeros estadounidenses". Reseñas de ciencias cuaternarias . 48 : 1–6. Código Bibliográfico : 2012QSRv ... 48 .... 1M . doi : 10.1016 / j.quascirev.2012.05.014 .
- ^ a b c d e Clague, John J .; Mathewes, Rolf W. y Ager, Thomas A. (2004). "Ambientes del noroeste de América del Norte antes del último máximo glacial" . En DB Madsen (ed.). Entrando en América: el noreste de Asia y Beringia antes del último máximo glacial . Prensa de la Universidad de Utah. ISBN 978-0-87480-786-8.
- ^ a b c d e f Vasil'ev, Sergey A .; Kuzmin, Yaroslav V .; Orlova, Lyubov A. y Dementiev, Vyacheslav N. (2002). "Cronología basada en radiocarbono del Paleolítico en Siberia y su relevancia para el poblamiento del Nuevo Mundo" . Radiocarbono . 44 (2): 503–530. doi : 10.1017 / s0033822200031878 .
- ↑ a b c Graf, Kelly E. (2009). "Colonización humana moderna de la estepa gigantesca: una vista desde el centro-sur de Siberia" (PDF) . En Marta Camps; Parth Chauhan (eds.). Libro de consulta de transiciones paleolíticas . Saltador. págs. 479–501. doi : 10.1007 / 978-0-387-76487-0_32 . ISBN 978-0-387-76478-8.
- ^ a b c d e Fedje, Daryl W .; Mackie, Quentin; Dixon, E. James y Heaton, Timothy H. (2004). "Medio ambiente de Wisconsin tardío y visibilidad arqueológica a lo largo de la costa noroeste del norte" . En DB Madsen (ed.). Entrando en América: el noreste de Asia y Beringia antes del último máximo glacial . Prensa de la Universidad de Utah. ISBN 978-0-87480-786-8.
- ↑ a b c d e Erlandson, Jon M. y Braje, Todd J. (2011). "¿De Asia a América en barco? Paleogeografía, paleoecología y puntas derivadas del Pacífico noroccidental". Cuaternario Internacional . 239 (1–2): 28–37. Código Bibliográfico : 2011QuInt.239 ... 28E . doi : 10.1016 / j.quaint.2011.02.030 .
- ↑ a b c Erlandson, Jon M .; Graham, Michael H .; Bourque, Bruce J .; et al. (2007). "La hipótesis de la carretera Kelp: ecología marina, la teoría de la migración costera y el poblamiento de las Américas". The Journal of Island and Coastal Archaeology . 2 (2): 161-174. doi : 10.1080 / 15564890701628612 . S2CID 140188874 .
- ^ a b c Vachula, RS; Huang, Y .; Russell, JM; et al. (20 de mayo de 2020). "Los biomarcadores sedimentarios reafirman los impactos humanos en los ecosistemas del norte de Beringia durante el último período glacial" . Boreas . 49 (3): 514-525. doi : 10.1111 / bor.12449 .
- ^ a b c Vachula, RS; Huang, Y .; Longo, WM; et al. (13 de diciembre de 2018). "La evidencia de los seres humanos de la Edad de Hielo en el este de Beringia sugiere una migración temprana a América del Norte". Reseñas de ciencias cuaternarias . 205 : 35–44. doi : 10.1016 / j.quascirev.2018.12.003 .
- ^ Blanco, Phillip M. (2006). Cronología de los indios americanos: cronologías del mosaico americano . Greenwood. pag. 1. ISBN 978-0-313-33820-5.
- ^ a b c d Wells, Spencer y Read, Mark (2002). El viaje del hombre: una odisea genética . Casa al azar. págs. 138–140. ISBN 978-0-8129-7146-0.
- ^ Lovgren, Stefan (13 de marzo de 2008). "Las Américas se asentaron hace 15.000 años, dice un estudio" . National Geographic .
- ↑ a b c d Bonatto, Sandro L. y Salzano, Francisco M. (1997). "Una única y temprana migración para el poblamiento de las Américas respaldada por datos de secuencia de ADN mitocondrial" . Actas de la Academia Nacional de Ciencias . 94 (5): 1866–1871. Código Bibliográfico : 1997PNAS ... 94.1866B . doi : 10.1073 / pnas.94.5.1866 . PMC 20009 . PMID 9050871 .
- ↑ a b Cinq-Mars, J. (1979). "Cueva de pez azul 1: un depósito de cueva de Beringio oriental del Pleistoceno tardío en el norte de Yukón". Revista canadiense de arqueología (3): 1–32. JSTOR 41102194 .
- ↑ a b Bonnichsen, Robson (1978). "Argumentos críticos para los artefactos del Pleistoceno de la cuenca del Viejo Cuervo, Yukon: una declaración preliminar" . En Alan L. Bryan (ed.). El hombre primitivo en América desde una perspectiva Circum-Pacific . Documentos ocasionales No. 1. Edmonton: Investigaciones arqueológicas Departamento Internacional de Antropología, Universidad de Alberta . págs. 102-118. ISBN 9780888649997.
- ^ a b c Oppenheimer, Stephen. "Viaje de la humanidad" . Fundación Bradshaw .
- ↑ a b c d Goodyear, Albert C. (2005). "Evidencia de sitios Pre-Clovis en el este de Estados Unidos" . En Robson Bonnichsen; et al. (eds.). Orígenes paleoamericanos: más allá de Clovis . Poblamiento de las Américas. Centro para el Estudio de los Primeros Americanos, Universidad A&M de Texas. págs. 103–112. ISBN 978-1-60344-812-3.
- ↑ a b c d Dillehay, Thomas (2000). El asentamiento de las Américas: una nueva prehistoria . Nueva York: Basic Books. ISBN 978-0-465-07669-7.
- ^ Kaplan, Sarah (24 de octubre de 2018). "Las puntas de lanza más antiguas del continente proporcionan nuevas pistas sobre los primeros estadounidenses" . Washington Post .
- ^ Pedersen, Mikkel W .; Ruter, Anthony; Schweger, Charles; et al. (10 de agosto de 2016). "Viabilidad postglacial y colonización en el corredor libre de hielo de América del Norte" . Naturaleza . 537 (7618): 45–49. Código bibliográfico : 2016Natur.537 ... 45P . doi : 10.1038 / nature19085 . PMID 27509852 . S2CID 4450936 .
- ^ Chung, Emily (August 10, 2016). "Popular theory on how humans populated North America can't be right, study shows: Ice-free corridor through Alberta, B.C. not usable by humans until after Clovis people arrived". CBC News. Retrieved August 10, 2016.
- ^ Figure 4 of Andrew, Kitchen (2008). "A Three-Stage Colonization Model for the Peopling of the Americas". PLOS ONE. 3 (2): e1596. Bibcode:2008PLoSO...3.1596K. doi:10.1371/journal.pone.0001596. PMC 2223069. PMID 18270583.
- ^ Morlan, Richard E. (March 4, 2015). "Old Crow Basin". The Canadian Encyclopedia. Historica Canada.
- ^ Bryant, Vaughn M., Jr. (1998). "Pre-Clovis". In Guy Gibbon; et al. (eds.). Archaeology of Prehistoric Native America: An Encyclopedia. Garland reference library of the humanities. 1537. pp. 682–683. ISBN 978-0-8153-0725-9.
- ^ Handwerk, Brian (22 July 2020). "Discovery in Mexican Cave May Drastically Change the Known Timeline of Humans' Arrival to the Americas". Smithsonian Magazine.
- ^ Santos, G.M; Bird, M.I; Parenti, F.; et al. (2003). "A revised chronology of the lowest occupation layer of Pedra Furada Rock Shelter, Piauı́, Brazil: The Pleistocene peopling of the Americas". Quaternary Science Reviews. 22 (21–22): 2303–2310. Bibcode:2003QSRv...22.2303S. doi:10.1016/S0277-3791(03)00205-1.
- ^ van Vark, G.N.; Kuizenga, D. & Williams, F.L. (June 2003). "Kennewick and Luzia: lessons from the European Upper Paleolithic". American Journal of Physical Anthropology. 121 (2): 181–184, discussion 185–188. doi:10.1002/ajpa.10176. PMID 12740961.
• Fiedel, Stuart J. (2004). "The Kennewick Follies: 'New' Theories about the Peopling of the Americas". Journal of Anthropological Research. 60 (1): 75–110. doi:10.1086/jar.60.1.3631009. JSTOR 3631009. S2CID 163722475.
• González-José, R .; Bortolini, MC; Santos, FR & Bonatto, SL (octubre de 2008). "El poblamiento de América: variación de la forma craneofacial a escala continental y su interpretación desde una mirada interdisciplinar" . Revista Estadounidense de Antropología Física . 137 (2): 175-187. doi : 10.1002 / ajpa.20854 . PMID 18481303 . S2CID 32748672 . - ↑ Moreno-Mayar, J. Víctor; Vinner, Lasse; de Barros Damgaard, Peter; de la Fuente, Constanza; et al. (7 de diciembre de 2018). "Primeras dispersiones humanas dentro de las Américas" . Ciencia . 362 (6419): eaav2621. Código Bibliográfico : 2018Sci ... 362.2621M . doi : 10.1126 / science.aav2621 . PMID 30409807 .
- ^ Posth, Cosimo; Nakatsuka, Nathan; Lazaridis, Iosif; Skoglund, Pontus; et al. (15 de noviembre de 2018). "Reconstrucción de la historia profunda de la población de América Central y del Sur" . Celular . 175 (5): 1185-1197.e22. doi : 10.1016 / j.cell.2018.10.027 . ISSN 0092-8674 . PMC 6327247 . PMID 30415837 .
- ^ a b c Adovasio, J. M; Donahue, J. & Stuckenrath, R. (1990). "The Meadowcroft Rockshelter Rasdiocarbon Chronology 1975–1990". American Antiquity. 55 (2): 348–354. doi:10.2307/281652. JSTOR 281652.
• Hirst, K. Kris (October 23, 2017). "What Does cal BP Mean?". Thoughtco.com. Retrieved October 30, 2018. - ^ Holen, Steven R .; Deméré, Thomas A .; Fisher, Daniel C .; et al. (2017). "Un sitio arqueológico de 130.000 años en el sur de California, Estados Unidos". Naturaleza . 544 (7651): 479–483. Bibcode : 2017Natur.544..479H . doi : 10.1038 / nature22065 . PMID 28447646 .
- ^ Rincon, Paul (26 April 2017). "First Americans claim sparks controversy". BBC News. Retrieved 30 April 2017.
Michael R. Waters commented that "To demonstrate such early occupation of the Americas requires the presence of unequivocal stone artifacts. There are no unequivocal stone tools associated with the bones... this site is likely just an interesting paleontological locality." Chris Stringerdijo que "las afirmaciones extraordinarias requieren evidencia extraordinaria; cada aspecto requiere el escrutinio más fuerte", y agregó que "deben haberse requerido fuerzas altas y concentradas para aplastar los huesos de mastodonte más gruesos, y el entorno de depósito de baja energía aparentemente no proporciona una alternativa obvia para los humanos que usan el adoquines pesados encontrados con los huesos.
- ^ Pitulko, VV; Nikolsky, PA; Girya, E. Yu; et al. (2 de enero de 2004). "El sitio de Yana RHS: humanos en el Ártico antes del último máximo glacial". Ciencia . 303 (5654): 52–56. Código Bibliográfico : 2004Sci ... 303 ... 52P . doi : 10.1126 / science.1085219 . ISSN 0036-8075 . PMID 14704419 . S2CID 206507352 .
- ^ a b c d e f g Tamm, Erika; Kivisild, Toomas; Reidla, Maere; et al. (2007). "Beringian Standstill and Spread of Native American Founders". PLOS ONE. 2 (9): e829. Bibcode:2007PLoSO...2..829T. doi:10.1371/journal.pone.0000829. PMC 1952074. PMID 17786201.
- ^ a b c d Cocina, Andrew; Miyamoto, Michal M. y Mulligan, Connie J. (2008). "Un modelo de colonización en tres etapas para el poblamiento de las Américas" . PLOS ONE . 3 (2): e1596. Código Bibliográfico : 2008PLoSO ... 3.1596K . doi : 10.1371 / journal.pone.0001596 . PMC 2223069 . PMID 18270583 .
- ^ Goebel, Ted y Buvit, Ian (2011). Del Yenisei al Yukón: interpretación de la variabilidad del ensamblaje lítico en Beringia del Pleistoceno tardío / Holoceno temprano . Centro para el estudio de los primeros estadounidenses, Texas A&M University Press. pag. 5. ISBN 978-1-60344-384-5.
- ↑ a b Skoglund, Pontus & Reich, David (diciembre de 2016). "Una visión genómica del poblamiento de las Américas" (PDF) . Opinión Actual en Genética y Desarrollo . 41 : 27–35. doi : 10.1016 / j.gde.2016.06.016 . PMC 5161672 . PMID 27507099 .
Recientemente, llevamos a cabo una prueba rigurosa de la hipótesis nula de una sola población fundadora de centroamericanos y sudamericanos utilizando datos de todo el genoma de diversos nativos americanos. Detectamos una señal estadísticamente clara que vincula a los nativos americanos de la región amazónica de Brasil con los australo-melanesios y los isleños de Andaman ('australasianos') actuales. Específicamente, encontramos que los australasianos comparten significativamente más variantes genéticas con algunas poblaciones amazónicas, incluidas las que hablan idiomas tupi, que con otros nativos americanos. Llamamos a este antiguo linaje de nativos americanos putativo "Población Y" en honor a Ypykuéra, que significa "antepasado" en la familia de lenguas Tupi.
- ↑ a b c Kemp, Brian M .; Malhi, Ripan S .; McDonough, John; et al. (2007). "Análisis genético de restos esqueléticos del Holoceno temprano de Alaska y sus implicaciones para el asentamiento de las Américas" (PDF) . Revista Estadounidense de Antropología Física . 132 (4): 605–621. CiteSeerX 10.1.1.576.7832 . doi : 10.1002 / ajpa.20543 . PMID 17243155 .
- ^ a b c d e f Perego, Ugo A .; Achilli, Alessandro; Angerhofer, normando; et al. (2009). "Rutas de migración paleoindias distintivas de Beringia marcadas por dos haplogrupos de ADNmt raros" . Biología actual . 19 (1): 1–8. doi : 10.1016 / j.cub.2008.11.058 . PMID 19135370 . S2CID 9729731 .
- ^ a b Derenko, Miroslava; Malyarchuk, Boris; Grzybowski, Tomasz; et al. (21 de diciembre de 2010). "Origen y dispersión post-glacial de los haplogrupos C y D de ADN mitocondrial en el norte de Asia" . PLOS ONE . 5 (12): e15214. Código Bibliográfico : 2010PLoSO ... 515214D . doi : 10.1371 / journal.pone.0015214 . PMC 3006427 . PMID 21203537 .
- ^ Bortolini, Maria-Catira; Salzano, Francisco M .; Thomas, Mark G .; et al. (2003). "Evidencia del cromosoma Y para diferentes historias demográficas antiguas en las Américas" (PDF) . Revista Estadounidense de Genética Humana . 73 (3): 524–539. doi : 10.1086 / 377588 . PMC 1180678 . PMID 12900798 .
- ↑ a b c Schurr, Theodore G. (mayo de 2000). "ADN mitocondrial y el poblamiento del nuevo mundo" (PDF) . Científico estadounidense . 88 (3): 246. Código Bibliográfico : 2000AmSci..88..246S . doi : 10.1511 / 2000.3.246 .
- ^ a b Zakharov, I.A.; Derenko, M.V.; Maliarchuk, B.A.; et al. (12 January 2006). "Mitochondrial DNA variation in the aboriginal populations of the Altai-Baikal region: implications for the genetic history of North Asia and America". Annals of the New York Academy of Sciences. 1011 (1): 21–35. Bibcode:2004NYASA1011...21Z. doi:10.1196/annals.1293.003. PMID 15126280. S2CID 37139929.
- ↑ a b c d e Starikovskaya, Elena B .; Sukernik, Rem I .; Derbeneva, Olga A .; et al. (Enero de 2005). "Diversidad de ADN mitocondrial en poblaciones indígenas de la extensión sur de Siberia y los orígenes de los haplogrupos nativos americanos" . Annals of Human Genetics . 69 (Parte 1): 67–89. doi : 10.1046 / j.1529-8817.2003.00127.x . PMC 3905771 . PMID 15638829 .
- ^ Sikora, Martin; Pitulko, Vladimir V.; Sousa, Vitor C.; et al. (2019). "The population history of northeastern Siberia since the Pleistocene" (PDF). Nature. 570 (7760): 182–188. Bibcode:2019Natur.570..182S. doi:10.1038/s41586-019-1279-z. PMID 31168093. S2CID 174809069.
- ^ a b c d Adachi, Noboru; Shinoda, Ken-ichi; Umetsu, Kazuo & Matsumura, Hirofumi (marzo de 2009). "Análisis de ADN mitocondrial de esqueletos de Jōmon del sitio Funadomari, Hokkaido, y su implicación para los orígenes de los nativos americanos". Revista Estadounidense de Antropología Física . 138 (3): 255-265. doi : 10.1002 / ajpa.20923 . PMID 18951391 .
- ^ Adachi, Noboru; Shinoda, Ken-ichi; Umetsu, Kazuo; et al. (Noviembre de 2011). "Análisis de ADN mitocondrial de esqueletos de Hokkaido Jōmon: restos de linajes maternos arcaicos en el borde suroeste de la ex Beringia". Revista Estadounidense de Antropología Física . 146 (3): 346–360. doi : 10.1002 / ajpa.21561 . PMID 21953438 .
- ^ Li, Hong-Chuan; Biggar, Robert J.; Miley, Wendell J.; et al. (2004). "Provirus load in breast milk and risk of mother-to-child transmission of Human T Lymphotropic Virus Type I". The Journal of Infectious Diseases. 190 (7): 1275–1278. doi:10.1086/423941. PMID 15346338.
- ^ a b c d Verdonck, K.; González, E.; Van Dooren, S.; et al. (April 2007). "Human T-lymphotropic virus 1: recent knowledge about an ancient infection". The Lancet Infectious Diseases. 7 (4): 266–281. doi:10.1016/S1473-3099(07)70081-6. PMID 17376384.
- ^ Gessain, A.; Gallo, R.C. & Franchini, G. (April 1992). "Low degree of human T-cell leukemia/lymphoma virus type I genetic drift in vivo as a means of monitoring viral transmission and movement of ancient human populations". Journal of Virology. 66 (4): 2288–2295. doi:10.1128/JVI.66.4.2288-2295.1992. PMC 289023. PMID 1548762.
- ^ Ishida, Takafumi; Yamamoto, Kohtaro; Omoto, Keiichi; et al. (Septiembre de 1985). "Prevalencia de un retrovirus humano en los nativos japoneses: evidencia de un posible origen antiguo". Revista de Infección . 11 (2): 153-157. doi : 10.1016 / s0163-4453 (85) 92099-7 . PMID 2997332 .
- ^ a b Miura, T.; Fukunaga, T.; Igarashi, T.; et al. (February 1994). "Phylogenetic subtypes of human T-lymphotropic virus type I and their relations to the anthropological background". Proceedings of the National Academy of Sciences of the United States of America. 91 (3): 1124–1127. Bibcode:1994PNAS...91.1124M. doi:10.1073/pnas.91.3.1124. PMC 521466. PMID 8302841.
- ^ Picard, FJ; Coulthart, MB; Oger, J .; et al. (Noviembre de 1995). "Virus linfotrópico T humano tipo 1 en nativos costeros de Columbia Británica: afinidades filogenéticas y posibles orígenes" . Revista de Virología . 69 (11): 7248–56. doi : 10.1128 / JVI.69.11.7248-7256.1995 . PMC 189647 . PMID 7474147 .
- ^ Li, Hong-Chuan; Fujiyoshi, Toshinobu; Lou, Hong; et al. (December 1999). "The presence of ancient human T-cell lymphotropic virus type I provirus DNA in an Andean mummy". Nature Medicine. 5 (12): 1428–1432. doi:10.1038/71006. PMID 10581088. S2CID 12893136.
- ↑ a b Coulthart, Michael B .; Posada, David; Crandall, Keith A. y Dekaband, Gregory A. (marzo de 2006). "Sobre la ubicación filogenética de las secuencias del virus de la leucemia de células T humanas tipo 1 asociadas a una momia andina" . Infección, Genética y Evolución . 6 (2): 91–96. doi : 10.1016 / j.meegid.2005.02.001 . PMC 1983367 . PMID 16503510 .
- ↑ a b Gonzaleza, Silvia; Huddart, David; Israde-Alcántara, Isabel; et al. (30 de marzo de 2015). "Sitios paleoindios de la cuenca de México: evidencia de estratigrafía, tefrocronología y datación" (PDF) . Cuaternario Internacional . 363 : 4-19. Código bibliográfico : 2015QuInt.363 .... 4G . doi : 10.1016 / j.quaint.2014.03.015 .
- ↑ a b González-José, Rolando; González-Martín, Antonio; Hernández, Miquel; et al. (4 de septiembre de 2003). "Evidencia craneométrica de supervivencia paleoamericana en Baja California". Naturaleza . 425 (6953): 62–65. Bibcode : 2003Natur.425 ... 62G . doi : 10.1038 / nature01816 . PMID 12955139 . S2CID 4423359 .
- ^ Dillehay, Thomas D. (4 September 2003). "Tracking the first Americans". Nature. 425 (6953): 23–24. doi:10.1038/425023a. PMID 12955120. S2CID 4421265.
- ^ Fiedel, Stuart J. (Spring 2004). "The Kennewick follies: "new" theories about the peopling of the Americas". Journal of Anthropological Research. 60 (1): 75–110. doi:10.1086/jar.60.1.3631009. JSTOR 3631009. S2CID 163722475.
- ^ Chatters, James C .; Kennett, Douglas J .; Asmerom, Yemane; et al. (16 de mayo de 2014). "Esqueleto humano del Pleistoceno tardío y mtDNA vinculan a los paleoamericanos y los nativos americanos modernos" (PDF) . Ciencia . 344 (6185): 750–754. Código bibliográfico : 2014Sci ... 344..750C . doi : 10.1126 / science.1252619 . PMID 24833392 . S2CID 206556297 . Archivado desde el original (PDF) el 13 de julio de 2015.
- ↑ de Azvedo, Soledad; Bortolini, Maria C .; Bonatto, Sandro L .; et al. (Enero de 2015). "Restos antiguos y primer poblamiento de las Américas: reevaluación del cráneo de Hoyo Negro". Revista Estadounidense de Antropología Física . 148 (3): 514-521. doi : 10.1002 / ajpa.22801 . PMID 26174009 .
• Azevedo, Soledad de; Quinto-Sánchez, Mirsha; Paschetta, Carolina & González-José, Rolando (28 February 2017). ""The first human settlement of the New World " A closer look at craniofacial variation and evolution of early and late Holocene Native American groups". Quaternary International. 431 (part B): 152–167. Bibcode:2017QuInt.431..152D. doi:10.1016/j.quaint.2015.11.012. - ^ Seong, Chuntaek (December 2008). "Tanged points, microblades and late paleolithic hunting in Korea". Antiquity. 82 (318): 871–883. doi:10.1017/s0003598x00097647.
- ^ Waters, Michael R. & Stafford, Thomas W. (23 February 2007). "Redefining the age of Clovis: implications for the peopling of the Americas". Science. 315 (5815): 1122–1126. Bibcode:2007Sci...315.1122W. doi:10.1126/science.1137166. PMID 17322060. S2CID 23205379.
- ^ Jenkins, Dennis L.; Davis, Loren G.; Stafford, Thomas W., Jr; et al. (13 July 2012). "Clovis Age Western Stemmed Projectile Points and Human Coprolites at the Paisley Caves". Science. 337 (6091): 223–228. Bibcode:2012Sci...337..223J. doi:10.1126/science.1218443. PMID 22798611. S2CID 40706795.
- ^ a b Vajda, Edward J. (18 April 2017). "Dene-Yeniseian". Oxford Bibliographies Online. doi:10.1093/OBO/9780199772810-0064.
- ^ Flegontov, Pavel; Altınışık, N. Ezgi; Changmai, Piya; et al. (13 de octubre de 2017). "Legado genético paleo-esquimal en América del Norte" . bioRxiv . doi : 10.1101 / 203018 . hdl : 21.11116 / 0000-0004-5D08-C . S2CID 90288469 .
• Flegontov, Pavel; Altınışık, N. Ezgi; Changmai, Piya; et al. (5 de junio de 2019). "Ascendencia genética paleo-esquimal y el poblamiento de Chukotka y América del Norte" (PDF) . Naturaleza . 570 (7760): 236–240. Código Bib : 2019Natur.570..236F . doi : 10.1038 / s41586-019-1251-y . ISSN 0028-0836 . PMC 6942545 . PMID 31168094 . - ^ Handwerk, Brian (12 de febrero de 2010). "Rostro de humano antiguo extraído del ADN del cabello; el genoma pinta la imagen del hombre de la cultura extinta de Groenlandia" . Noticias de National Geographic .
- ^ Fladmark, Knute R. (enero de 1979). "Rutas: corredores de migración alternativos para el hombre primitivo en América del Norte". Antigüedad americana . 44 (1): 55–69. doi : 10.2307 / 279189 . JSTOR 279189 .
- ^ Callaway, Ewen (11 de agosto de 2016). "El ADN de plantas y animales sugiere que los primeros estadounidenses tomaron la ruta costera" . Naturaleza . 536 (7615): 138. Bibcode : 2016Natur.536..138C . doi : 10.1038 / 536138a . PMID 27510205 .
- ^ Summer, Thomas (10 August 2016). "Humans may have taken different path into Americas than thought Arctic passage wouldn't have provided enough food for the earliest Americans' journey". Science News.
- ^ Rasmussen, Morten; Anzick, Sarah L.; Waters, Michael R.; Skoglund, Pontus; DeGiorgio, Michael; Stafford, Thomas W., Jr; et al. (February 2014). "The genome of a Late Pleistocene human from aClovis burial site in western Montana". Nature. 506 (7487): 225–229. Bibcode:2014Natur.506..225R. doi:10.1038/nature13025. PMC 4878442. PMID 24522598.
- ^ Gornitz, Vivian (January 2007). "Sea Level Rise, After the Ice Melted and Today". Goddard Institute for Space Studies. NASA. Archived from the original on 2007-02-02. Retrieved 23 April 2015.
- ^ Hetherington, Renée; Barrie, J. Vaughn; MacLeod, Roger & Wilson, Michael (February 2004). "Quest for the Lost Land". Geotimes.
- ^ "California islands give up evidence of early seafaring: Numerous artifacts found at late Pleistocene sites on the Channel Islands". Science Daily. University of Oregon. 3 March 2011.
Bibliografía
- Bradley, Bruce y Stanford, Dennis J. (2004). "El corredor del borde del hielo del Atlántico Norte: una posible ruta del Paleolítico al Nuevo Mundo". Arqueología mundial . 36 (4): 459–478. CiteSeerX 10.1.1.694.6801 . doi : 10.1080 / 0043824042000303656 . S2CID 161534521 .
- Bradley, Bruce y Stanford, Dennis J. (2006). "La conexión Solutrean-Clovis: respuesta a Straus, Meltzer y Goebel". Arqueología mundial . 38 (4): 704–714. doi : 10.1080 / 00438240601022001 . JSTOR 40024066 . S2CID 162205534 .
- Stanford, Dennis J .; Bradley, Bruce (2012). Primeros americanos pre-Clovis: El origen de la cultura Clovis de Estados Unidos . Prensa de la Universidad de California. ISBN 978-0-520-22783-5.
- Stanford, Dennis J. y Bradley, Bruce A. (2013). Al otro lado del hielo del Atlántico: el origen de la cultura Clovis de Estados Unidos . Prensa de la Universidad de California. ISBN 978-0-520-27578-2.
- Dixon, E. James (1993). Quest for the Origins of the First Americans. University of New Mexico. ISBN 978-0-8263-1406-2.
- Dixon, E. James (1999). Bones, Boats & Bison: Archeology and the First Colonization of Western North America. University of New Mexico Press. ISBN 978-0-8263-2138-1.
- Erlandson, Jon M. (2013). Early Hunter-Gatherers of the California Coast. Springer Science & Business Media. ISBN 978-1-4757-5042-3.
- Erlandson, Jon M. (2001). "The Archaeology of Aquatic Adaptations: Paradigms for a New Millennium". Journal of Archaeological Research. 9 (4): 287–350. doi:10.1023/a:1013062712695. S2CID 11120840.
- Erlandson, Jon M. (2002). "Anatomically modern humans, maritime voyaging, and the Pleistocene colonization of the Americas". In Nina G. Jablonski (ed.). The First Americans: The Pleistocene Colonization of the New World. California Academy of Sciences. pp. 59–92. ISBN 978-0-940228-50-4.
- Erlandson, Jon. M.; Graham, M. H.; Bourque, Bruce J.; et al. (30 October 2007). "The Kelp Highway Hypothesis: Marine Ecology, The Coastal Migration Theory, and the Peopling of the Americas". Journal of Island and Coastal Archaeology. 2 (2): 161–174. doi:10.1080/15564890701628612. S2CID 140188874.
- Eshleman, Jason A .; Malhi, Ripan S. y Glenn Smith, David (2003). "Estudios de ADN mitocondrial de los nativos americanos: conceptos y conceptos erróneos de la prehistoria de la población de las Américas" . Antropología evolutiva . 12 (1): 7–18. doi : 10.1002 / evan.10048 . S2CID 17049337 .
- Fedje, Daryl W. y Christensen, Tina (octubre de 1999). "Modelado de Paleoshorelines y localización de sitios costeros del Holoceno temprano en Haida Gwaii". Antigüedad americana . 64 (4): 635–652. doi : 10.2307 / 2694209 . JSTOR 2694209 .CS1 maint: date and year (link)
- Greenman, EF (febrero de 1963). "El Paleolítico Superior y el Nuevo Mundo". Antropología actual . 4 (1): 41–66. doi : 10.1086 / 200337 . JSTOR 2739818 . S2CID 144250630 .
- Hola, Jody (25 de mayo de 2005). "Sobre el número de fundadores del nuevo mundo: un retrato genético de la población de los pueblos de las Américas" . PLOS Biología . 3 (6): e193. doi : 10.1371 / journal.pbio.0030193 . PMC 1131883 . PMID 15898833 .
- Jablonski, Nina G. (2002). Los primeros estadounidenses: la colonización del Pleistoceno del Nuevo Mundo . Academia de Ciencias de California . ISBN 978-0-940228-50-4.
- Jones, Peter N. (2005). Respeto por los antepasados: afiliación cultural indígena americana en el oeste americano . Instituto Bauu. ISBN 978-0-9721349-2-7.
- Korotayev, Andrey; Berezkin, Yuri E .; Borinskaya, Svetlana A .; Davletshin, Albert I .; Khaltourina, Daria A. (2017). "¿Qué genes y mitos trajeron las diferentes oleadas de poblamiento de América al Nuevo Mundo?" . En Leonid E. Grinin; Andrey V. Korotayev; Yuri E. Berezkin (eds.). Historia y Matemáticas: Economía, Demografía, Cultura y Civilizaciones Cósmicas . págs. 9–77. ISBN 978-5-7057-5247-8.
- Lauber, Patricia (2003). Quién vino primero: nuevas pistas para los estadounidenses prehistóricos . Libros para niños de National Geographic Soc. ISBN 978-0-7922-8228-0.
- Matson, RG y Coupland, Gary (2016). La prehistoria de la costa noroeste . Taylor y Francis. ISBN 978-1-315-41739-4.
- Meltzer, David J. (2009). Primeros pueblos en un mundo nuevo: colonización de América de la Edad de Hielo . Prensa de la Universidad de California. ISBN 978-0-520-94315-5.
- Nieve, Dean R. (1996). "Los primeros estadounidenses y la diferenciación de culturas de cazadores-recolectores" . En Bruce G. Trigger; Wilcomb E. Washburn (eds.). La historia de Cambridge de los pueblos nativos de las Américas: América del Norte . Volumen 1: Parte 1. Cambridge University Press. págs. 125–199. ISBN 978-0-521-57392-4.
|volume=has extra text (help) - Wells, Spencer (2002). El viaje del hombre: una odisea genética . Prensa de la Universidad de Princeton. ISBN 0-691-11532-X.
Enlaces externos
- The Paleoindian Database – The University of Tennessee, Department of Anthropology.
- "The first Americans: How and when were the Americas populated?", Earth, January 2016
- "When Did Humans Come to the Americas?" – Smithsonian Magazine February 2013
- The Paleoindian Period – United States Department of the Interior, National Park Service
- Shepard Krech III, Paleoindians and the Great Pleistocene Die-Off – American Academy of Arts and Sciences, National Humanities Center, 2008.
- Journey of Man: A Genetic Odyssey (movie) on YouTube – by Spencer Wells – PBS and National Geographic Channel, 2003 – 120 Minutes, UPC/EAN: 841887001267
| vte History of the Americas | ||
|---|---|---|
| History |
|  |
| Settlement |
| |
| Sociedades |
| |
| Relacionada |
| |
| Liza |
| |
| Cronología |
| |
| ||
| vtmiPueblos indígenas de las Américas | |||||
|---|---|---|---|---|---|
| Prehistoria |
|  | |||
| Mitología / Religión |
| ||||
| Colonización europea |
| ||||
| Grupos modernos por país |
| ||||
| Temas relacionados |
| ||||
| |||||
| vtmi América del Norte precolombina | |
|---|---|
| |
| Archaeological cultures |
|
| Archaeologicalsites | 
|
| Humanremains | |
| Miscellaneous |
|
| |