| KCNMA1 | |||||||
|---|---|---|---|---|---|---|---|
 La estructura de dominio de los canales BK | |||||||
| Identificadores | |||||||
| Símbolo | KCNMA1 | ||||||
| Alt. simbolos | SLO | ||||||
| Gen NCBI | 3778 | ||||||
| HGNC | 6284 | ||||||
| OMIM | 600150 | ||||||
| RefSeq | NM_002247 | ||||||
| UniProt | Q12791 | ||||||
| Otros datos | |||||||
| Lugar | Chr. 10 q22 | ||||||
| |||||||
{kind=link}
| KCNMB1 | |||||||
|---|---|---|---|---|---|---|---|
| Identificadores | |||||||
| Símbolo | KCNMB1 | ||||||
| Gen NCBI | 3779 | ||||||
| HGNC | 6285 | ||||||
| OMIM | 603951 | ||||||
| RefSeq | NM_004137 | ||||||
| UniProt | Q16558 | ||||||
| Otros datos | |||||||
| Lugar | Chr. 5 q34 | ||||||
| |||||||
| KCNMB2 | |||||||
|---|---|---|---|---|---|---|---|
| Identificadores | |||||||
| Símbolo | KCNMB2 | ||||||
| Gen NCBI | 10242 | ||||||
| HGNC | 6286 | ||||||
| OMIM | 605214 | ||||||
| RefSeq | NM_181361 | ||||||
| UniProt | Q9Y691 | ||||||
| Otros datos | |||||||
| Lugar | Chr. 3 q26.32 | ||||||
| |||||||
| KCNMB3 | |||||||
|---|---|---|---|---|---|---|---|
| Identificadores | |||||||
| Símbolo | KCNMB3 | ||||||
| Alt. simbolos | KCNMB2, KCNMBL | ||||||
| Gen NCBI | 27094 | ||||||
| HGNC | 6287 | ||||||
| OMIM | 605222 | ||||||
| RefSeq | NM_171828 | ||||||
| UniProt | Q9NPA1 | ||||||
| Otros datos | |||||||
| Lugar | Chr. 3 q26.3-q27 | ||||||
| |||||||
| KCNMB3L | |
|---|---|
| Identificadores | |
| Símbolo | KCNMB3L |
| Alt. simbolos | KCNMB2L, KCNMBLP |
| Gen NCBI | 27093 |
| HGNC | 6288 |
| RefSeq | NG_002679 |
| Otros datos | |
| Lugar | Chr. 22 q11.1 |
| KCNMB4 | |||||||
|---|---|---|---|---|---|---|---|
| Identificadores | |||||||
| Símbolo | KCNMB4 | ||||||
| Gen NCBI | 27345 | ||||||
| HGNC | 6289 | ||||||
| OMIM | 605223 | ||||||
| RefSeq | NM_014505 | ||||||
| UniProt | Q86W47 | ||||||
| Otros datos | |||||||
| Lugar | Chr. 12 q15 | ||||||
| |||||||
| Subunidad alfa del canal de potasio BK activado por calcio | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identificadores | |||||||||
| Símbolo | BK_channel_a | ||||||||
| Pfam | PF03493 | ||||||||
| InterPro | IPR003929 | ||||||||
| |||||||||
Los canales BK (potasio grande) son canales de potasio activados por calcio de gran conductancia , [1] también conocidos como Maxi-K , slo1 o Kca1.1 . Los canales BK son canales de potasio activados por voltaje que conducen grandes cantidades de iones de potasio (K + ) a través de la membrana celular , de ahí su nombre, potasio grande . Estos canales pueden activarse (abrirse) por medios eléctricos o aumentando las concentraciones de Ca 2+ en la célula. [2] [3] Los canales BK ayudan a regular los procesos fisiológicos, comoritmos de comportamiento circadianos y excitabilidad neuronal. [4] Los canales BK también están involucrados en muchos procesos en el cuerpo, ya que es un canal ubicuo. Tienen una estructura tetramérica que se compone de un dominio transmembrana , un dominio sensor de voltaje, un dominio del canal de potasio y un dominio C-terminal citoplasmático , con muchas estructuras de rayos X como referencia. Su función es repolarizar el potencial de membrana permitiendo que el potasio fluya hacia afuera, en respuesta a una despolarización o aumento de los niveles de calcio.
Estructura [ editar ]
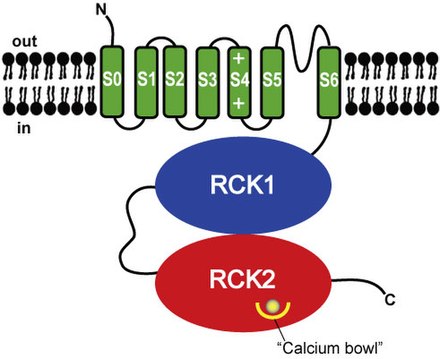
Estructuralmente, los canales BK son homólogos a los canales de potasio activados por voltaje y ligando , y tienen un sensor de voltaje y un poro como dominio que atraviesa la membrana y un dominio citosólico para la unión de calcio y magnesio intracelulares . [5] Cada monómero de la subunidad alfa formadora de canales es el producto del gen KCNMA1 (también conocido como Slo1). La subunidad Slo1 tiene tres dominios estructurales principales, cada uno con una función distinta: el dominio de detección de voltaje (VSD) detecta el potencial de membranaa través de la membrana, el dominio citosólico (detecta la concentración de calcio, iones Ca²⁺) y el dominio de puerta de poros (PGD) que se abre y se cierra para regular la permeación de potasio . La puerta de activación reside en el PGD, que se encuentra en el lado citosólico de S6 o en el filtro de selectividad (la selectividad es la preferencia de un canal para conducir un ión específico). [5] El dominio de detección de voltaje y el dominio controlado por poros se denominan colectivamente dominios que atraviesan la membrana y están formados por los segmentos transmembrana S1-S4 y S5-S6, respectivamente. Dentro de la hélice S4 contiene una serie de residuos cargados positivamente que sirven como sensor de voltaje primario . [6]
Los canales BK son bastante similares a los canales K⁺ activados por voltaje, sin embargo, en los canales BK solo un residuo cargado positivamente (Arg213) está involucrado en la detección de voltaje a través de la membrana. [5] También es exclusivo de los canales BK un segmento S0 adicional, este segmento es necesario para la modulación de la subunidad β . [7] [8] y sensibilidad al voltaje. [9]
El dominio citosólico está compuesto por dos dominios RCK (regulador de la conductancia de potasio), RCK1 y RCK2. Estos dominios contienen dos sitios de unión de Ca²⁺ de alta afinidad : uno en el dominio RCK1 y el otro en una región denominada cuenco de Ca²⁺ que consta de una serie de residuos de ácido aspártico (Asp) que se encuentran en el dominio RCK2. El sitio de unión de Mg²⁺ está ubicado entre el VSD y el dominio citosólico, que está formado por: residuos Asp dentro del bucle S0-S1, residuos de asparagina en el extremo citosólico de S2 y residuos de glutamina en RCK1. [5]Al formar el sitio de unión de Mg²⁺, dos residuos provienen del RCK1 de una subunidad Slo1 y los otros dos residuos provienen del VSD de la subunidad vecina. Para que estos residuos coordinen el ion Mg²⁺, el VSD y el dominio citosólico de las subunidades vecinas deben estar muy cerca. [5] Las subunidades beta moduladoras (codificadas por KCNMB1 , KCNMB2 , KCNMB3 o KCNMB4 ) pueden asociarse con el canal tetramérico . Hay cuatro tipos de subunidades β (β1-4), cada una de las cuales tiene diferentes patrones de expresión que modifican las propiedades de activación del canal BK. La subunidad β1 es la principal responsable de las células del músculo liso.expresión, las subunidades β2 y β3 se expresan neuronalmente, mientras que β4 se expresa dentro del cerebro . [5] El VSD se asocia con el PGD a través de tres interacciones principales:
- Conexión física entre el VSD y el PGD a través del enlazador S4-S5.
- Interacciones entre el enlazador S4-S5 y el lado citosólico de S6.
- Interacciones entre S4 y S5 de una subunidad vecina.
Reglamento [ editar ]
Los canales BK están asociados y modulados por una amplia variedad de factores intra y extracelulares, como subunidades auxiliares (β, γ), Slobs (proteína de unión a slo), fosforilación , voltaje de membrana , ligandos químicos (Ca²⁺, Mg²⁺), PKC , Las subunidades α de BK se ensamblan 1: 1 con cuatro tipos auxiliares diferentes de subunidades β (β1, β2, β3 o β4). [10]
Se ha encontrado que el tráfico y la expresión de los canales BK en la membrana plasmática están regulados por distintos motivos de corte y empalme ubicados dentro de los dominios RCK intracelulares C-terminales. En particular, una variante de corte y empalme que excluía estos motivos impidió la expresión de los canales BK en la superficie celular y sugiere que tal mecanismo impacta en la fisiología y fisiopatología . [10]
Los canales de BK en el sistema vascular están modulados por agentes producidos naturalmente en el cuerpo, como la angiotensina II (Ang II), glucosa alta o ácido araquidónico (AA) que se modula en la diabetes por estrés oxidativo (ROS). [10]
Una sensibilidad de voltaje más débil permite que los canales BK funcionen en una amplia gama de potenciales de membrana. Esto asegura que el canal pueda realizar correctamente su función fisiológica. [11]
La inhibición de la actividad del canal BK por fosforilación de S695 por la proteína quinasa C (PKC) depende de la fosforilación de S1151 en el extremo C de la subunidad alfa del canal. Sólo es necesario que se produzca una de estas fosforilaciones en la estructura tetramérica para que la inhibición sea satisfactoria. La proteína fosfatasa 1 contrarresta la fosforilación de S695. PKC reduce la probabilidad de apertura del canal acortando el tiempo de apertura del canal y prolongando el estado cerrado del canal. La PKC no afecta la conductancia de un solo canal, la dependencia del voltaje o la sensibilidad al calcio de los canales BK. [11]
Mecanismo de activación [ editar ]
Los canales BK se activan sinérgicamente mediante la unión de iones de calcio y magnesio , pero también se pueden activar mediante la dependencia del voltaje. [10] La activación dependiente de Ca²⁺ ocurre cuando el Ca²⁺ intracelular se une a dos sitios de unión de alta afinidad : uno ubicado en el extremo C-terminal del dominio RCK2 (cuenco Ca²⁺) y el otro ubicado en el dominio RCK1. [5] El sitio de unión dentro del dominio RCK1 tiene una afinidad algo menor por el calcio que el cuenco de Ca²⁺, pero es responsable de una mayor porción de la sensibilidad al Ca²⁺. [12] El voltaje y el calcio activan los canales BK usando dos mecanismos paralelos, con los sensores de voltaje.y los sitios de unión de Ca²⁺ se acoplan a la puerta de activación de forma independiente, excepto por una interacción débil entre los dos mecanismos. El recipiente de Ca²⁺ acelera la cinética de activación a concentraciones bajas de Ca²⁺, mientras que el sitio RCK1 influye tanto en la cinética de activación como en la de desactivación. [11] Un modelo de mecanismo fue propuesto originalmente por Monod, Wyman y Changeux, conocido como modelo MWC. El modelo MWC para los canales BK explica que un cambio conformacional de la puerta de activación en la apertura del canal se acompaña de un cambio conformacional en el sitio de unión de Ca²⁺, lo que aumenta la afinidad de la unión de Ca²⁺. [12]
La activación dependiente de magnesio de los canales BK se activa a través de un sitio de unión a metales de baja afinidad que es independiente de la activación dependiente de Ca²⁺. El sensor de Mg²⁺ activa los canales BK cambiando el voltaje de activación a un rango más negativo. Mg²⁺ activa el canal solo cuando el dominio del sensor de voltaje permanece en el estado activado. El dominio de la cola citosólica (CTD) es un sensor químico que tiene múltiples sitios de unión para diferentes ligandos . El CTD activa el canal BK cuando se une con Mg²⁺ intracelular para permitir la interacción con el dominio del sensor de voltaje (VSD). [11] El magnesio está coordinado predominantemente por seis átomos de oxígeno de las cadenas laterales de los residuos que contienen oxígeno, los grupos carbonilo de la cadena principal.en proteínas o moléculas de agua . [12] D99 en el extremo C-terminal del bucle S0-S1 y N172 en el bucle S2-S3 contienen oxígenos de cadena lateral en el dominio del sensor de voltaje que son esenciales para la unión de Mg²⁺. Al igual que el modelo de activación dependiente de Ca²⁺, la activación dependiente de Mg²⁺ también puede describirse mediante un modelo de activación de MCW alostérico. Mientras que el calcio activa el canal en gran medida independientemente del sensor de voltaje, el magnesio activa canal por canal mediante una interacción electrostática con el sensor de voltaje. [12] Esto también se conoce como el modelo Nudging, en el que el magnesio activa el canal presionando el sensor de voltaje a través de interacciones electrostáticas e involucra las interacciones entrecadenas laterales en diferentes dominios estructurales. [5] La energía proporcionada por la unión de voltaje, Ca²⁺ y Mg²⁺ se propagará a la puerta de activación de los canales BK para iniciar la conducción de iones a través del poro. [5]
Efectos sobre la neurona, el órgano y el cuerpo en su conjunto [ editar ]
Nivel celular [ editar ]
Los canales BK ayudan a regular tanto la activación de neuronas como la liberación de neurotransmisores . [13] Esta modulación de la transmisión sináptica y la descarga eléctrica a nivel celular se debe a la expresión del canal BK junto con otros canales de potasio-calcio. [10] La apertura de estos canales provoca un impulso hacia el potencial de equilibrio de potasio y, por lo tanto, juega un papel en la aceleración de la repolarización de los potenciales de acción . [10] Esto permitiría efectivamente una estimulación más rápida. [10] También se juega un papel en la configuración de la repolarización general de las células y, por lo tanto,después de la hiperpolarización (AHP) de los potenciales de acción. [14] El papel que tienen los canales BK en la fase rápida de AHP se ha estudiado ampliamente en el hipocampo. [14] También puede desempeñar un papel en la inhibición de la liberación de neurotransmisores. [15] Hay muchos canales BK en las células de Purkinje del cerebelo , lo que destaca su papel en la coordinación y función motora . [14] Además, los canales BK desempeñan un papel en la modulación de la actividad de las dendritas , así como de los astrocitos y la microglía . [15] No solo juegan un papel en el SNC (sistema nervioso central ) sino también en las contracciones del músculo liso , la secreción de células endocrinas y la proliferación de células. [13] Varias subunidades γ durante el desarrollo temprano del cerebro están involucradas en la excitabilidad neuronal y en las células no excitables a menudo son responsables como fuerza impulsora del calcio. [10] Por lo tanto, estas subunidades pueden ser objetivos de tratamientos terapéuticos como activadores del canal BK. [10] Hay más evidencia de que la inhibición de los canales BK evitaría la salida de potasio y, por lo tanto, reduciría el uso de ATP , permitiendo de hecho la supervivencia neuronal en entornos con poco oxígeno. [10]Los canales BK también pueden funcionar como un protector neuronal en términos tales como limitar la entrada de calcio en las células a través de la oxidación de metionina . [10]
Nivel de órgano [ editar ]
Los canales BK también juegan un papel importante en la audición . [14] Esto se encontró cuando la subunidad ɑ de BK se eliminó en ratones y se observó la pérdida progresiva de las células ciliadas cocleares y, por lo tanto, la pérdida de audición. [14] Los canales BK no solo participan en la audición, sino también en los ritmos circadianos . Las proteínas de unión a Slo (Slobs) pueden modular los canales BK en función de los ritmos circadianos en las neuronas. [10] Los canales BK se expresan en el núcleo supraquiasmático (SCN), que se caracteriza por influir en la fisiopatología del sueño. [14] Los abridores de canal BK también pueden tener un efecto protector en elsistema cardiovascular . [10] A una concentración baja de calcio, los canales BK tienen un mayor impacto en el tono vascular . [10] Además, el sistema de señalización de los canales BK en el sistema cardiovascular influye en el funcionamiento del flujo sanguíneo coronario . [10] Una de las funciones de la subunidad β en el cerebro incluye la inhibición de los canales BK, lo que permite la ralentización de las propiedades del canal, así como la capacidad de ayudar en la prevención de convulsiones en el lóbulo temporal . [10]
Nivel de función corporal [ editar ]
Las mutaciones de los canales BK, que dan como resultado una menor cantidad de expresión en el ARNm , son más comunes en personas con discapacidades mentales (por hipofunción [15] ), esquizofrénicas o autistas . [10] Además, el aumento de la repolarización causado por mutaciones del canal BK puede conducir a la dependencia del alcohol al inicio de discinesias , epilepsia o trastornos del movimiento paroxístico. [10] Los canales BK no solo son importantes en muchos procesos celulares en el adulto, sino que también son cruciales para el suministro de una nutrición adecuada al feto en desarrollo . [10] Por lo tanto,el estrógeno puede provocar un aumento de la densidad de los canales BK en el útero . [10] Sin embargo, se ha encontrado una mayor expresión de los canales BK en las células tumorales , y esto podría influir en la futura terapia contra el cáncer , que se analiza más en la sección de farmacología. [10] Los canales BK son omnipresentes en todo el cuerpo y, por lo tanto, tienen un gran impacto en el cuerpo como un todo y a un nivel más celular, como se discutió.
Farmacología [ editar ]
Posibles problemas [ editar ]
Surgen varios problemas cuando hay un déficit en los canales BK. Las consecuencias del mal funcionamiento del canal BK pueden afectar el funcionamiento de una persona de muchas maneras, algunas más potencialmente mortales que otras. Los canales BK pueden ser activados por contaminantes exógenos y gasotransmisores endógenos monóxido de carbono , [16] [17] óxido nítrico y sulfuro de hidrógeno. [18] Las mutaciones en las proteínas involucradas con los canales BK o genes que codifican los canales BK están involucradas en muchas enfermedades. Un mal funcionamiento de los canales BK puede proliferar en muchos trastornos como: epilepsia , cáncer , diabetes , asma e hipertensión.. [13] Específicamente, el defecto β1 puede aumentar la presión arterial y la retención de hidrosalina en el riñón . [13] Se ha descubierto que tanto la pérdida de función como la ganancia de mutaciones de función están involucradas en trastornos como la epilepsia y el dolor crónico . [15] Además, los aumentos en la activación del canal BK, a través de mutantes de ganancia de función y amplificación, tiene vínculos con la epilepsia y el cáncer. [13] Además, los canales BK desempeñan un papel tanto en los tumores como en los cánceres. En ciertos cánceres, se puede encontrar gBK, una variante del canal iónico llamado canal BK del glioma. [14] Se sabe que los canales BK influyen de alguna manera en la división de las células durante la replicación., que cuando no se regula puede provocar cánceres y tumores. [14] Además, un aspecto estudiado incluye la migración de las células cancerosas y el papel en el que los canales BK pueden facilitar esta migración, aunque todavía se desconoce mucho. [14] Otra razón por la que la comprensión del canal BK es importante tiene que ver con su papel en la cirugía de trasplante de órganos . Esto se debe a la activación de los canales BK que influyen en la repolarización del potencial de membrana en reposo . [10] Por lo tanto, la comprensión es crucial para la seguridad de un trasplante eficaz.
Desarrollos actuales [ editar ]
Los canales BK se pueden utilizar como dianas farmacológicas para el tratamiento de varios trastornos médicos, incluido el accidente cerebrovascular [19] y la vejiga hiperactiva . [20] Ha habido intentos de desarrollar moléculas sintéticas dirigidas a los canales BK, [21] sin embargo, sus esfuerzos han demostrado ser en gran medida ineficaces hasta ahora. Por ejemplo, BMS-204352, una molécula desarrollada por Bristol-Myers Squibb , no logró mejorar el resultado clínico en pacientes con accidente cerebrovascular en comparación con el placebo . [22] Sin embargo, ha habido cierto éxito del agonista de los canales BKCa, BMS-204352, en el tratamiento de los déficits observados en Fmr1. ratones knockout , un modelo del síndrome de X frágil . [23] [24] Los canales BK también funcionan como bloqueadores en la isquemia y son un foco en la investigación de su uso como terapia para el accidente cerebrovascular. [10]
Direcciones futuras [ editar ]
Existen muchas aplicaciones para las estrategias terapéuticas que involucran canales BK. Ha habido investigaciones que muestran que un bloqueo de los canales BK da como resultado un aumento en la liberación de neurotransmisores, lo que indica de manera efectiva las posibilidades terapéuticas futuras en la mejora de la cognición , la memoria mejorada y el alivio de la depresión . [13] Una respuesta conductual al alcohol también está modulada por los canales BK, [10] por lo tanto, una mayor comprensión de esta relación puede ayudar al tratamiento en pacientes alcohólicos . El estrés oxidativo en los canales BK puede conducir a los deterioros negativos que la reducción de la presión arterial a través de la relajación cardiovascular tiene sobre el envejecimiento y la enfermedad.[10] Por lo tanto, el sistema de señalización puede participar en el tratamiento de la hipertensión y la aterosclerosis [10] mediante el direccionamiento de la subunidad ɑ para prevenir estos efectos perjudiciales. Además, el papel conocido que pueden desempeñar los canales BK en el cáncer y los tumores es limitado. Por lo tanto, no hay mucho conocimiento actual sobre aspectos específicos de los canales BK que pueden influir en tumores y cánceres. [14] Es fundamental realizar más estudios, ya que esto podría conducir a un inmenso desarrollo en los tratamientos para quienes padecen cáncer y tumores. Se sabe que las epilepsias se deben a la sobreexcitabilidad de las neuronas, cuyos canales BK tienen un gran impacto en el control de la hiperexcitabilidad. [4]Por tanto, la comprensión podría influir en el tratamiento de la epilepsia. En general, los canales BK son un objetivo para futuros agentes farmacológicos que pueden usarse para tratamientos benévolos de enfermedades.
Ver también [ editar ]
- Subunidad alfa-1 del canal de potasio activado por calcio
- Canal de potasio activado por calcio
- Canal de potasio dependiente de voltaje
Referencias [ editar ]
- ^ Zang K, Zhang Y, Hu J, Wang Y (2018). "El canal de potasio (BK) activado por voltaje y calcio de gran conductancia y la epilepsia". Objetivos de fármacos para trastornos neurológicos y del SNC . 17 (4): 248-254. doi : 10.2174 / 1871527317666180404104055 . PMID 29623857 .
- ^ Miller, C. (2000). Biología del genoma, 1 (4), reviews0004.1. https://dx.doi.org/10.1186/gb-2000-1-4-reviews0004
- ^ Yuan, P., Leonetti, M., Pico, A., Hsiung, Y. y MacKinnon, R. (2010). Estructura del aparato de activación de Ca2 + del canal BK humano a una resolución de 3,0 A. Science, 329 (5988), 182-186. https://dx.doi.org/10.1126/science.1190414
- ↑ a b N'Gouemo P (noviembre de 2011). "Dirigirse a los canales BK (potasio grande) en la epilepsia" . Opinión de expertos sobre objetivos terapéuticos . 15 (11): 1283–95. doi : 10.1517 / 14728222.2011.620607 . PMC 3219529 . PMID 21923633 .
- ^ a b c d e f g h i Lee US, Cui J (septiembre de 2010). "Activación del canal BK: conocimientos estructurales y funcionales" . Tendencias en neurociencias . 33 (9): 415-23. doi : 10.1016 / j.tins.2010.06.004 . PMC 2929326 . PMID 20663573 .
- ^ Atkinson NS, Robertson GA, Ganetzky B (agosto de 1991). "Un componente de los canales de potasio activados por calcio codificados por el locus slo de Drosophila". Ciencia . 253 (5019): 551–5. doi : 10.1126 / science.1857984 . PMID 1857984 . S2CID 11317087 .
- ^ Morrow JP, Zakharov SI, Liu G, Yang L, Sok AJ, Marx SO (marzo de 2006). "Definición de los dominios del canal BK necesarios para la modulación de la subunidad beta1" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 103 (13): 5096–101. doi : 10.1073 / pnas.0600907103 . PMC 1458800 . PMID 16549765 .
- ^ Wallner M, Meera P, Toro L (diciembre de 1996). "Determinante para la regulación de la subunidad beta en canales de K + sensibles a voltaje de alta conductancia y Ca (2 +) - sensibles: una región transmembrana adicional en el extremo N" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 93 (25): 14922–7. doi : 10.1073 / pnas.93.25.14922 . PMC 26238 . PMID 8962157 .
- ^ Koval OM, Fan Y, Rothberg BS (marzo de 2007). "Un papel para el segmento transmembrana S0 en la compuerta dependiente del voltaje de los canales BK" . La Revista de Fisiología General . 129 (3): 209-20. doi : 10.1085 / jgp.200609662 . PMC 2151615 . PMID 17296928 .
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z Hermann A, Sitdikova GF, Weiger TM (agosto de 2015). "Estrés oxidativo y canales de potasio activado por calcio máximo (BK)" . Biomoléculas . 5 (3): 1870–911. doi : 10.3390 / biom5031870 . PMC 4598779 . PMID 26287261 .
- ↑ a b c d Yang H, Zhang G, Cui J (2015). "Canales BK: múltiples sensores, una puerta de activación" . Fronteras en fisiología . 6 : 29. doi : 10.3389 / fphys.2015.00029 . PMC 4319557 . PMID 25705194 .
- ↑ a b c d Cui J, Yang H, Lee US (marzo de 2009). "Mecanismos moleculares de activación del canal BK" . Ciencias de la vida celular y molecular . 66 (5): 852–75. doi : 10.1007 / s00018-008-8609-x . PMC 2694844 . PMID 19099186 .
- ^ a b c d e f Yu M, Liu SL, Sun PB, Pan H, Tian CL, Zhang LH (enero de 2016). "Toxinas peptídicas y bloqueadores de moléculas pequeñas de los canales BK" . Acta Pharmacologica Sinica . 37 (1): 56–66. doi : 10.1038 / aps.2015.139 . PMC 4722972 . PMID 26725735 .
- ↑ a b c d e f g h i j Bentzen BH, Olesen SP, Rønn LC, Grunnet M (2014). "Activadores del canal BK y sus perspectivas terapéuticas" . Fronteras en fisiología . 5 : 389. doi : 10.3389 / fphys.2014.00389 . PMC 4191079 . PMID 25346695 .
- ↑ a b c d Contet C, Goulding SP, Kuljis DA, Barth AL (2016). "Canales BK en el sistema nervioso central". Big on Bk - Información actual sobre la función de los canales de K + activados por voltaje y Ca2 + de gran conductancia en los niveles molecular, celular y sistémico . Revista Internacional de Neurobiología . 128 . págs. 281–342. doi : 10.1016 / bs.irn.2016.04.001 . ISBN 9780128036198. PMC 4902275 . PMID 27238267 .
- ^ Dubuis E, Potier M, Wang R, Vandier C (febrero de 2005). "La inhalación continua de monóxido de carbono atenúa el desarrollo de hipertensión pulmonar hipóxica presumiblemente a través de la activación de los canales de BKCa" . Investigación cardiovascular . 65 (3): 751–61. doi : 10.1016 / j.cardiores.2004.11.007 . PMID 15664403 .
- ^ Hou S, Xu R, Heinemann SH, Hoshi T (marzo de 2008). "El sensor de Ca2 + de alta afinidad RCK1 confiere sensibilidad al monóxido de carbono a los canales Slo1 BK" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 105 (10): 4039–43. doi : 10.1073 / pnas.0800304105 . PMC 2268785 . PMID 18316727 .
- ^ Sitdikova GF, Weiger TM, Hermann A (febrero de 2010). "El sulfuro de hidrógeno aumenta la actividad del canal de potasio activado por calcio (BK) de las células tumorales pituitarias de rata". Pflügers Archiv . 459 (3): 389–97. doi : 10.1007 / s00424-009-0737-0 . PMID 19802723 . S2CID 23073556 .
- ^ Gribkoff VK, Starrett JE, Dworetzky SI (abril de 2001). "Canales de potasio Maxi-K: forma, función y modulación de una clase de reguladores endógenos de calcio intracelular". El neurocientífico . 7 (2): 166–77. doi : 10.1177 / 107385840100700211 . PMID 11496927 . S2CID 8791803 .
- ^ Layne JJ, Nausch B, Olesen SP, Nelson MT (febrero de 2010). "La activación del canal BK por NS11021 disminuye la excitabilidad y la contractilidad del músculo liso de la vejiga urinaria" . Revista estadounidense de fisiología. Fisiología reguladora, integradora y comparada . 298 (2): R378–84. doi : 10.1152 / ajpregu.00458.2009 . PMC 2828174 . PMID 19923353 .
- ^ Gribkoff VK, Winquist RJ (mayo de 2005). "Moduladores de canales de cationes activados por voltaje para el tratamiento del accidente cerebrovascular". Opinión de expertos sobre fármacos en investigación . 14 (5): 579–92. doi : 10.1517 / 13543784.14.5.579 . PMID 15926865 . S2CID 10236998 .
- ^ Jensen BS (2002). "BMS-204352: un abridor de canales de potasio desarrollado para el tratamiento del accidente cerebrovascular" . Revisiones de medicamentos del SNC . 8 (4): 353–60. doi : 10.1111 / j.1527-3458.2002.tb00233.x . PMC 6741660 . PMID 12481191 .
- ^ Laumonnier F, Roger S, Guérin P, Molinari F, M'rad R, Cahard D, Belhadj A, Halayem M, Persico AM, Elia M, Romano V, Holbert S, Andres C, Chaabouni H, Colleaux L, Constant J , Le Guennec JY, Briault S (2006). "Asociación de un déficit funcional del canal BKCa, un regulador sináptico de la excitabilidad neuronal, con autismo y retraso mental". La Revista Estadounidense de Psiquiatría . 163 (9): 1622–1629. doi : 10.1176 / ajp.2006.163.9.1622 . PMID 16946189 . S2CID 25225269 .
- ^ Hébert B; Pietropaolo S; Memes; Laudier B; Laugeray A; Doisne N; Quartier A; Lefeuvre S; Tengo L; Cahard D; Laumonnier F; Crusio WE ; Pichon J; Menuet A; Perche O; Briault S (2014). "Rescate de fenotipos del síndrome X frágil en ratones Fmr1 KO por una molécula abridor de canal BKCa" . Revista Orphanet de Enfermedades Raras . 9 : 124. doi : 10.1186 / s13023-014-0124-6 . PMC 4237919 . PMID 25079250 .
Lectura adicional [ editar ]
- Ge L, Hoa NT, Wilson Z, Arismendi-Morillo G, Kong XT, Tajhya RB, Beeton C, Jadus MR (octubre de 2014). "Grandes canales de iones de potasio (BK) en biología, enfermedad y posibles dianas para la inmunoterapia del cáncer" . Inmunofarmacología internacional . 22 (2): 427–43. doi : 10.1016 / j.intimp.2014.06.040 . PMC 5472047 . PMID 25027630 .
- Kyle BD, Braun AP (2014). "La regulación de la actividad del canal BK por modificaciones pre y postraduccionales" . Fronteras en fisiología . 5 : 316. doi : 10.3389 / fphys.2014.00316 . PMC 4141542 . PMID 25202279 .
- Nardi A, Olesen SP (2008). "Moduladores de canal BK: una descripción completa". Química Medicinal Actual . 15 (11): 1126–46. doi : 10.2174 / 092986708784221412 . PMID 18473808 .
- Zhang J, Yan J (2014). "Regulación de canales BK por subunidades γ auxiliares" . Fronteras en fisiología . 5 : 401. doi : 10.3389 / fphys.2014.00401 . PMC 4197896 . PMID 25360119 .
Enlaces externos [ editar ]
- Canales BK + en los encabezados de temas médicos (MeSH) de la Biblioteca Nacional de Medicina de EE. UU .
- "Canales de potasio activados por calcio" . Base de datos IUPHAR de receptores y canales de iones . Unión Internacional de Farmacología Básica y Clínica.