| Digenea | |
|---|---|
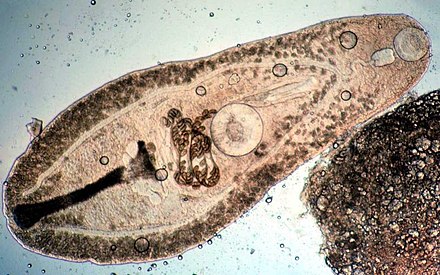 | |
| Helicometra sp. ( Plagiorchiida : Opecoelidae ) del intestino de una llama Cardinal pescado | |
| clasificación cientifica | |
| Reino: | Animalia |
| Filo: | Platelmintos |
| Clase: | Rhabditophora |
| Clade : | Bothrioneodermata |
| Superorden: | Neodermata |
| Clade : | Trematoda |
| Clade : | Digenea Carus , 1863 |
| Familias | |
{kind=link}
Digenea (Gr. Dis - double, Genos - raza) es una clase de trematodos en el filo Platyhelminthes , que consiste en gusanos planos parásitos (conocidos como trematodos ) con un tegumento sincitial y, generalmente, dos ventosas , una ventral y una oral. Los adultos suelen vivir dentro del tracto digestivo, pero se encuentran en todos los sistemas de órganos de todas las clases de vertebrados . Alguna vez se pensó que estaban relacionados con la Monogenea , ahora se reconoce que están más cerca de la Aspidogastrea y que los Monogenea están más estrechamente aliados con los Cestoda . Hasta la fecha se han descrito alrededor de 6.000 especies.
Morfología [ editar ]
Funciones clave [ editar ]
Los rasgos característicos de la Digenea incluyen un tegumento sincitial; es decir, un tegumento donde las uniones entre las células se rompen y un solo citoplasma continuo rodea a todo el animal. Un tegumento similar se encuentra en otros miembros de los Neodermata ; un grupo de platelmintos que comprende Digenea, Aspidogastrea , Monogenea y Cestoda . Los digeneos poseen un plan corporal vermiforme , no segmentado y tienen un parénquima sólido sin cavidad corporal ( celoma ) como en todos los platelmintos.
Por lo general, hay dos ventosas , una ventosa oral anterior que rodea la boca y una ventosa ventral a veces denominada acetábulo , en la superficie ventral. La ventosa oral rodea la boca, mientras que la ventral es un órgano muscular ciego sin conexión con ninguna estructura interna.
Un monostoma es un gusano con una ventosa (oral). Los trematodos con una ventosa oral y un acetábulo en el extremo posterior del cuerpo se denominan anfistomos . Los distomes son trematodos con una ventosa oral y una ventral ventral, pero la ventosa ventral está en otro lugar que no sea la posterior. Estos términos son comunes en la literatura más antigua, cuando se pensaba que reflejaban relaciones sistemáticas dentro de los grupos. Han caído en desuso en la taxonomía digénica moderna.
Sistema reproductivo [ editar ]
La gran mayoría de los digeneos son hermafroditas . Es probable que esto sea una adaptación a la baja abundancia dentro de los huéspedes, lo que permite que el ciclo de vida continúe cuando solo un individuo infecta con éxito al huésped final. La fertilización es interna, y los espermatozoides se transfieren a través del cirro al canal de Laurer o apertura genital . Un grupo clave de digeneos que son dioicos son los esquistosomas . La reproducción asexual en la primera etapa larvaria es ubicua.
Si bien la formación sexual de los huevos digeneanos y la reproducción asexual en la primera etapa larvaria (miracidium) se informa ampliamente, la biología del desarrollo de las etapas asexuales sigue siendo un problema. Los estudios de microscopía electrónica han demostrado que las bolas germinales visibles al microscopio óptico consisten en células que se dividen mitóticamente y que dan lugar a embriones y a una línea de nuevas células germinales que se incluyen en estas etapas embrionarias. Dado que la ausencia de procesos meióticos no está probada, la definición exacta sigue siendo dudosa.
Órganos masculinos [ editar ]
Protandry es la regla general entre los Digenea. Por lo general, hay dos testículos , pero algunos trematodos pueden tener más de 100. También están presentes los conductos eferentes , los conductos deferentes , la vesícula seminal , el conducto eyaculador y un cirro (análogo a un pene) generalmente (pero no siempre) encerrado en un saco cirro. . Los cirros pueden o no estar cubiertos de espinas proteicas. La conformación exacta de estos órganos dentro de los genitales terminales masculinos es taxonómicamente importante a nivel familiar y genérico.
Órganos femeninos [ editar ]
Por lo general, hay un solo ovario con un oviducto , un receptáculo seminal , un par de glándulas vitelinas (involucradas en la producción de yema y cáscara de huevo) con conductos, el ootipo (una cámara donde se forman los huevos), una colección compleja de células de glándulas llamadas Glándula de Mehlis , que se cree que lubrica el útero para el paso de los óvulos.
Además, algunos digeneos poseen un canal llamado Laurer's Canal , que va desde el oviducto hasta la superficie dorsal del cuerpo. La función de este canal es objeto de debate, pero puede utilizarse para inseminación en algunas especies o para la eliminación de productos de desecho de la reproducción en otras especies. La mayoría de los trematodos poseen un ovicapt, una porción agrandada del oviducto donde se une al ovario. Probablemente controla la liberación de óvulos y espacia su descenso por el útero.
El útero generalmente se abre en un atrio genital común que también recibió el órgano copulador masculino distal (cirro) antes de abrirse inmediatamente a la superficie externa del gusano. La parte distal del útero puede expandirse en un metratérmino, separada del útero proximal por un esfínter muscular, o puede estar revestida de espinas, como en los Monorchiidae y algunas otras familias.
Sistema digestivo [ editar ]
En la edad adulta, la mayoría de los digeneos poseen una boca terminal o subterminal, una faringe muscular que proporciona la fuerza para ingerir alimentos y un sistema digestivo ciego y bifurcado que consta de dos sacos tubulares llamados ciegos (sing. Ciego ). En algunas especies, los dos ciegos intestinales se unen posteriormente para formar un intestino en forma de anillo o ciclocele . En otros, el ciego puede fusionarse con la pared del cuerpo posteriormente para formar uno o más anos , o con la vesícula excretora para formar un uroprocto . Los digeneos también son capaces de absorber directamente los nutrientes a través del tegumento por pinocitosis y fagocitosis por el sincitium . La mayoría de los digeneos adultos ocurren en vertebrados.canal alimentario o sus órganos asociados , donde con mayor frecuencia pastan en el contenido de la luz (p. ej., alimentos ingeridos por el huésped, bilis, moco), pero también pueden alimentarse a través de la pared mucosa (p. ej., submucosa , sangre del huésped). Los trematodos sanguíneos, como los esquistosomas, espirorquidos y sanguinicólidos, se alimentan exclusivamente de sangre. Los estadios asexuales en los hospedadores intermediarios de moluscos se alimentan principalmente por absorción directa, aunque el estadio de redia que se encuentra en algunos grupos tiene boca, faringe e intestino simple y puede consumir activamente tejido del hospedador o incluso otros parásitos. Los estadios metacercariales enquistados y los estadios cercariales de vida libre no se alimentan.
Sistema nervioso [ editar ]
Los ganglios emparejados en el extremo anterior del cuerpo sirven como cerebro . A partir de este, los nervios se extienden hacia delante y hacia atrás. Los receptores sensoriales son, en su mayor parte, deficientes entre los adultos, aunque tienen células tangoreceptoras . Los estadios larvarios tienen muchos tipos de receptores sensoriales, incluidos los receptores de luz y los quimiorreceptores. La quimiorrecepción juega un papel importante en el reconocimiento y la localización de la larva milagrosa de vida libre de su huésped.
Ciclos de vida [ editar ]
Existe una desconcertante variedad de variaciones en el complejo ciclo de vida digeneano, y la plasticidad de este rasgo es probablemente la clave del éxito del grupo. En general, los ciclos de vida pueden tener dos, tres o cuatro huéspedes obligados (necesarios), a veces con hospedadores de transporte o paraténicos en el medio. El ciclo de vida de tres huéspedes es probablemente el más común. En casi todas las especies, el primer hospedador del ciclo de vida es un molusco . [2] Esto ha llevado a la inferencia de que el digenean ancestral era un parásito de moluscos y que posteriormente se agregaron hospedadores vertebrados.
La alternancia de generaciones sexuales y asexuales es una característica importante de los digeneos. Este fenómeno implica la presencia de varias generaciones discretas en un ciclo de vida.
Un ciclo de vida típico de un trematodo digeneo es el siguiente. Los huevos dejan al huésped vertebrado en las heces y utilizan diversas estrategias para infectar al primer huésped intermedio , en el que no se produce la reproducción sexual. Los digeneos pueden infectar al primer huésped intermedio (generalmente un caracol ) por medios pasivos o activos. Los huevos de algunos digeneos, por ejemplo, son comidos (pasivamente) por caracoles (o, raramente, por un gusano anélido ), [2] en los que proceden a eclosionar. Alternativamente, los huevos pueden incubar en el agua para liberar una larva ciliada que nada activamente, el miracidio , que debe ubicarse y penetrar la pared del cuerpo del caracol huésped.
Después de la eclosión o la penetración del caracol después de la ingestión, el miracidium se metamorfosea en un esporocisto madre simple en forma de saco . El esporociste madre se somete a una ronda de reproducción asexual interna , dando lugar a rediae (sing. Redia) o esporoquistes hijas . La segunda generación es, por tanto, la secuencia hija parthenita. Éstos, a su vez, experimentan una reproducción asexual adicional, lo que finalmente produce un gran número de la segunda etapa de vida libre, la cercaria (pl. Cercariae).
Las cercarias que nadan libremente dejan al huésped del caracol y se mueven a través del ambiente acuático o marino , a menudo usando una cola en forma de látigo, aunque se observa una tremenda diversidad de morfología de la cola. Las cercarias son infecciosas para el segundo hospedador en el ciclo de vida y la infección puede ocurrir de forma pasiva (p. Ej., Un pez consume una cercaria) o activamente (la cercaria penetra en el pez).
Los ciclos de vida de algunos digeneos incluyen solo dos huéspedes, el segundo es un vertebrado. En estos grupos, la madurez sexual se produce después de que la cercaria penetra en el segundo hospedador, que en este caso también es el hospedador definitivo . Los ciclos de vida de dos huéspedes pueden ser primarios (nunca hubo un tercer huésped) como en los Bivesiculidae , o secundarios (en un momento de la historia evolutiva hubo un tercer huésped, pero se ha perdido).
En los ciclos de vida de tres huéspedes, las cercarias se desarrollan en el segundo huésped intermedio hasta una etapa de reposo, la metacercaria , que generalmente está enquistada en un quiste de origen del huésped y del parásito, o encapsulada en una capa de tejido derivada únicamente del huésped. Esta etapa es infecciosa para el hospedador definitivo . La transmisión ocurre cuando el huésped definitivo se alimenta de un segundo huésped intermedio infectado. Las metacercarias se exquistan en el intestino del hospedador definitivo en respuesta a una variedad de señales físicas y químicas, como los niveles de pH intestinal , enzimas digestivas , temperatura , etc. Una vez excitados, los digeneos adultos migran a sitios más o menos específicos en el hospedador definitivo y en la vida. el ciclo se repite.
Evolución [ editar ]
Los orígenes evolutivos de Digenea se han debatido durante algún tiempo, pero parece que existe un acuerdo general en que el proto-digenean era un parásito de un molusco, posiblemente de la cavidad del manto. La evidencia de esto proviene de la ubicuidad de los moluscos como primeros huéspedes intermediarios de los digeneos y del hecho de que la mayoría de los aspidogastreanos (el grupo hermano de Digenea) también tienen asociaciones con moluscos. Se cree que los primeros trematodos (el nombre colectivo para digeneos y aspidogastreanos) probablemente evolucionaron a partir de rabdoceleros turbelosos que colonizaron la cavidad abierta del manto de los primeros moluscos.
Es probable que los ciclos de vida más complejos evolucionaron a través de un proceso de adición terminal, por el cual los digeneos sobrevivieron a la depredación de su molusco hospedador, probablemente por un pez. Se agregaron otros hosts mediante el mismo proceso hasta que se desarrolló la desconcertante diversidad moderna de patrones de ciclo de vida.
Familias importantes [ editar ]
Digenea incluye alrededor de 80 familias. [3] Se enumeran a continuación, organizados por orden.
Digenea
- Diplostomida
- Suborden Diplostomata
- Superfamilia Brachylaimoidea Joyeux & Foley, 1930
- Brachylaimidae Joyeux y Foley, 1930
- Leucochloridiidae Poche, 1907
- Superfamilia Diplostomoidea Poirier, 1886
- Lobo Brauninidae , 1903
- Cyathocotylidae Mühling, 1898
- Diplostomidae Poirier, 1886
- Proterodiplostomidae Dubois, 1936
- Strigeidae Railliet, 1919
- Superfamilia Schistosomatoidea Stiles & Hassall, 1898
- Aporocotylidae Odhner, 1912
- Schistosomatidae Stiles y Hassall, 1898
- Spirorchiidae Stunkard, 1921
- Superfamilia Brachylaimoidea Joyeux & Foley, 1930
- Suborden Diplostomata
- Plagiorchiida
- Apocreadiata
- Apocreadioidea Skrjabin, 1942
- Apocreadiidae Skrjabin, 1942
- Apocreadioidea Skrjabin, 1942
- Bivesiculata
- Bivesiculoidea
- Bivesiculidae Yamaguti, 1934
- Bivesiculoidea
- Bucephalata
- Bucephaloidea Poche, 1907
- Bucephalidae Poche, 1907
- Nuitrematidae Kurochkin, 1975
- Gymnophalloidea Odhner, 1905
- Botulisaccidae Yamaguti, 1971
- Fellodistomidae Nicoll, 1909
- Gymnophallidae Odhner, 1905
- Tandanicolidae Johnston, 1927
- Bucephaloidea Poche, 1907
- Equinostomata
- Echinostomatoidea Looss, 1902
- Calycodidae Dollfus, 1929
- Cyclocoelidae Stossich, 1902
- Echinochasmidae Odhner, 1910
- Echinostomatidae Looss, 1899
- Eucotylidae Cohn, 1904
- Fasciolidae Railliet, 1895
- Himasthlidae Odhner, 1910
- Philophthalmidae Looss, 1899
- Psilostomidae Looss, 1900
- Rhytidodidae Odhner, 1926
- Typhlocoelidae Harrah, 1922
- Echinostomatoidea Looss, 1902
- Haplosplanchnata
- Haplosplanchnoidea Poche, 1925
- Haplosplanchnidae Poche, 1926
- Haplosplanchnoidea Poche, 1925
- Hemiurata
- Azygioidea Lühe, 1909
- Azygiidae Lühe, 1909
- Hemiuroidea Looss, 1899
- Accacoeliidae Odhner, 1911
- Bathcotylidae Dollfus, 1932
- Bathycotylidae Dollfus, 1932
- Derogenidae Nicoll, 1910
- Dictysarcidae Skrjabin y Guschanskaja, 1955
- Didymozoidae Monticelli, 1888
- Gonocercidae Skrjabin y Guschanskaja, 1955
- Hemiuridae Looss, 1899
- Hirudinellidae Dollfus, 1932
- Isoparorchiidae Travassos, 1922
- Lecithasteridae Odhner, 1905
- Ptychogonimidae Dollfus, 1937
- Sclerodistomidae Odhner, 1927
- Sclerodistomoididae Gibson y Bray, 1979
- Syncoeliidae Looss, 1899
- Azygioidea Lühe, 1909
- Heronimata
- Barrio Heronimoidea , 1918
- Sala Heronimidae , 1918
- Barrio Heronimoidea , 1918
- Lepocreadiata
- Lepocreadioidea Odhner, 1905
- Aephnidiogenidae Yamaguti, 1934
- Deropristidae Cable y Hunninen, 1942
- Enenteridae Yamaguti, 1958
- Gorgocephalidae Manter, 1966
- Gyliauchenidae Fukui, 1929
- Lepidapedidae Yamaguti, 1958
- Lepocreadiidae Odhner, 1905
- Liliatrematidae Gubanov, 1953
- Lepocreadioidea Odhner, 1905
- Monorchiata
- Monorchioidea Odhner, 1911
- Lissorchiidae Magath, 1917
- Monorchiidae Odhner, 1911
- Monorchioidea Odhner, 1911
- Opisthorchiata
- Opisthorchioidea Braun, 1901
- Distrito de Cryptogonimidae , 1917
- Heterophyidae Leiper, 1909
- Opisthorchiidae Looss, 1899
- Opisthorchioidea Braun, 1901
- Pronocephalata
- Paramphistomoidea Fischoeder, 1901
- Cladorchiidae Fischoeder, 1901
- Mesometridae Poche, 1926
- Microscaphidiidae Looss, 1900
- Paramphistomidae Fischoeder, 1901
- Pronocephaloidea Looss, 1899
- Labicolidae Blair, 1979
- Notocotylidae Lühe, 1909
- Nudacotylidae Barker, 1916
- Opisthotrematidae Poche, 1926
- Pronocephalidae Looss, 1899
- Rhabdiopoeidae Poche, 1926
- Paramphistomoidea Fischoeder, 1901
- Transversotremata
- Transversotrematoidea Witenberg, 1944
- Transversotrematidae Witenberg, 1944
- Transversotrematoidea Witenberg, 1944
- Xiphidiata
- Allocreadioidea Looss, 1902
- Acanthocolpidae Lühe, 1906
- Allocreadiidae Looss, 1902
- Batrachotrematidae Dollfus y Williams, 1966
- Brachycladiidae Odhner, 1905
- Opecoelidae Ozaki, 1925
- Gorgoderoidea Looss, 1901
- Callodistomidae Odhner, 1910
- Dicrocoeliidae Looss, 1899
- Gorgoderidae Looss, 1899
- Haploporoidea Nicoll, 1914
- Atractotrematidae Yamaguti, 1939
- Haploporidae Nicoll, 1914
- Distrito de Microphalloidea , 1901
- Diplangidae Yamaguti, 1971
- Exotidendriidae Mehra, 1935
- Faustulidae Poche, 1926
- Sala de Microphallidae , 1901
- Pachypsolidae Yamaguti, 1958
- Phaneropsolidae Mehra, 1935
- Pleurogenidae Looss, 1899
- Prosthogonimidae Lühe, 1909
- Renicolidae Dollfus, 1939
- Zoogonidae Odhner, 1902
- Plagiorchioidea Lühe, 1901
- Auridistomidae Lühe, 1901
- Brachycoeliidae Looss, 1899
- Cephalogonimidae Looss, 1899
- Choanocotylidae Jue Sue y Platt, 1998
- Echinoporidae Krasnolobova y Timofeeva, 1965
- Encyclometridae Mehra, 1931
- Leptophallidae Dayal, 1938
- Macroderoididae McMullen, 1937
- Meristocotylidae Fischthal y Kuntz, 1981
- Ocadiatrematidae Fischthal y Kuntz, 1981
- Orientocreadiidae Yamaguti, 1958
- Plagiorchiidae Lühe, 1901
- Styphlotrematidae Baer, 1924
- Telorchiidae Looss, 1899
- Thrinascotrematidae Jue Sue y Platt, 1999
- Urotrematidae Poche, 1926
- Allocreadioidea Looss, 1902
- Apocreadiata
Infecciones humanas digénicas [ editar ]
Solo alrededor de 12 de las 6.000 especies conocidas son infecciosas para los humanos, pero algunas de estas especies son enfermedades importantes que afectan a más de 200 millones de personas. Las especies que infectan a los humanos se pueden dividir en grupos, los esquistosomas y los no esquistosomas.
Esquistosomas [ editar ]
Los esquistosomas se encuentran en el sistema circulatorio del hospedador definitivo. Los seres humanos se infectan después de que las cercarias que nadan libremente liberadas de los caracoles infectados penetran en la piel. Estos gusanos dioicos son largos y delgados, con un tamaño que varía de 10 a 30 mm de longitud a 0,2 a 1,0 mm de diámetro. Los machos adultos son más cortos y más gruesos que las hembras, y tienen un surco largo a lo largo de un lado del cuerpo en el que se sujeta la hembra. Las hembras alcanzan la madurez sexual después de haberse unido a un macho. Después de aparearse, los dos permanecen encerrados juntos por el resto de sus vidas. Pueden vivir varios años y producir muchos miles de huevos.
Las cuatro especies de esquistosomas que infectan a los humanos son miembros del género Schistosoma .
| Nombre científico | Primer anfitrión intermedio | Área endémica |
|---|---|---|
| Schistosoma mansoni | Biomphalaria spp. | África , América del Sur , Caribe , Medio Oriente |
| Schistosoma haematobium | Bulinus spp. | África , Medio Oriente |
| Schistosoma japonicum | Oncomelania spp. | China , Asia oriental , Filipinas |
| Schistosoma intercalatum | Bulinus spp. | África |
No esquistosomas [ editar ]
Las siete especies principales de no esquistosomas que infectan a los seres humanos se enumeran a continuación. Las personas se infectan después de ingerir quistes metacercariales en plantas o en carne de animales poco cocida. La mayoría de las especies habitan en el tracto gastrointestinal humano, donde arrojan huevos junto con las heces del huésped. Paragonimus westermani , que coloniza los pulmones, también puede pasar sus huevos en la saliva . Estos trematodos generalmente causan una patología leve en humanos, pero también pueden ocurrir efectos más graves.
| Nombre científico | Primer anfitrión intermedio | Modo de infección humana | Área endémica |
|---|---|---|---|
| Fasciolopsis buski | Segmentina sp. | Plantas | Asia , India |
| Heterophyes heterophyes | Pirinella [ cita requerida ] | Salmonete , Tilapia | Asia , Europa del Este , Egipto , Medio Oriente |
| Metagonimus yokogawaii | Semisulcospira sp. [ cita requerida ] | Carpa , Trucha | Siberia |
| Gastrodiscoides hominis | Helicorbis sp. [ cita requerida ] | Plantas | India , Vietnam , Filipinas |
| Clonorchis sinensis | Bulinus sp. | Pez | Este de Asia , América del Norte |
| Fasciola hepática | Galba truncatula | Plantas | Mundial |
| Paragonimus westermani | Oncomelania sp. | Cangrejos , cangrejos de río | Asia |
Referencias [ editar ]
- ^ Bray, RA .; Justine, JL. (2014). "Una revisión de Zoogonidae (Digenea: Microphalloidea) de peces de las aguas alrededor de Nueva Caledonia, con la descripción de Overstreetia cribbi n. Sp" . PeerJ . 2 : e292. doi : 10.7717 / peerj.292 . PMC 3961169 . PMID 24688868 .
- ^ a b "Principios del parasitismo: Digenea" . www.biology.ualberta.ca . Consultado el 9 de enero de 2020 .
- ^ Olson PD, Cribb TH, Tkach VV, Bray RA y Littlewood DTJ (2003). "Filogenia y clasificación de la Digenea (Platyhelminthes: Trematoda)". Revista Internacional de Parasitología 33 (7): 733–755. doi : 10.1016 / S0020-7519 (03) 00049-3
Notas [ editar ]
- Gibson, DI, Jones, A. y Bray, RA (2002). Key to the Trematoda, vol.1 ISBN 0-85199-547-0
- Littlewood DTJ y Bray RA (2001) Interrelationship of the Platyhelminthes. ISBN 0-7484-0903-3
- Yamaguti, S. (1971). Sinopsis de trematodos digenetic de vertebrados. Keigaku Publishing Co., Tokio.
Enlaces externos [ editar ]
| Wikimedia Commons tiene medios relacionados con Digenea . |
- Grupo de investigación sobre esquistosomas de la Universidad de Cambridge
- Gusanos parásitos en el Museo de Historia Natural de Londres
- Fishdisease.net