
La contracción muscular es la activación de sitios que generan tensión dentro de las células musculares . [1] [2] En fisiología , la contracción muscular no significa necesariamente un acortamiento muscular porque la tensión muscular se puede producir sin cambios en la longitud del músculo, como cuando se sostiene un libro pesado o una mancuerna en la misma posición. [1] La terminación de la contracción muscular es seguida por la relajación muscular , que es un retorno de las fibras musculares a su estado de generación de baja tensión. [1]
Las contracciones musculares se pueden describir en función de dos variables: longitud y tensión. [1] Una contracción muscular se describe como isométrica si la tensión muscular cambia pero la longitud del músculo permanece igual. [1] [3] [4] [5] En contraste, una contracción muscular es isotónica si la tensión muscular permanece igual durante toda la contracción. [1] [3] [4] [5] Si la longitud del músculo se acorta, la contracción es concéntrica; [1] [6] si la longitud del músculo se alarga, la contracción es excéntrica. En movimientos naturales que subyacen a la actividad locomotora., las contracciones musculares son multifacéticas ya que pueden producir cambios en la longitud y la tensión de forma variable en el tiempo. [7] Por lo tanto, es probable que ni la longitud ni la tensión permanezcan iguales en los músculos que se contraen durante la actividad locomotora.
En los vertebrados , las contracciones del músculo esquelético son neurogénicas ya que requieren información sináptica de las neuronas motoras . Una sola motoneurona puede inervar múltiples fibras musculares, lo que hace que las fibras se contraigan al mismo tiempo. Una vez inervados, los filamentos de proteína dentro de cada fibra del músculo esquelético se deslizan entre sí para producir una contracción, que se explica por la teoría del filamento deslizante . La contracción producida puede describirse como una contracción , sumatoria o tétanos, dependiendo de la frecuencia de los potenciales de acción.. En los músculos esqueléticos, la tensión muscular es máxima cuando el músculo se estira a una longitud intermedia como se describe en la relación longitud-tensión.
A diferencia del músculo esquelético, las contracciones de los músculos lisos y cardíacos son miogénicas (lo que significa que son iniciadas por las propias células del músculo liso o cardíaco en lugar de ser estimuladas por un evento externo como la estimulación nerviosa), aunque pueden ser moduladas por estímulos del sistema nervioso autónomo . Los mecanismos de contracción en estos tejidos musculares son similares a los de los tejidos del músculo esquelético.
Tipos

Las contracciones musculares se pueden describir en función de dos variables: fuerza y longitud. La fuerza misma se puede diferenciar como tensión o carga. La tensión muscular es la fuerza que ejerce el músculo sobre un objeto, mientras que una carga es la fuerza que ejerce un objeto sobre el músculo. [1] Cuando la tensión muscular cambia sin ningún cambio correspondiente en la longitud del músculo, la contracción del músculo se describe como isométrica. [1] [3] [4] [5] Si la longitud del músculo cambia mientras la tensión del músculo permanece igual, entonces la contracción del músculo es isotónica. [1] [3] [4] [5] En una contracción isotónica, la longitud del músculo puede acortarse para producir una contracción concéntrica o alargarse para producir una contracción excéntrica.[1] [6] En los movimientos naturales que subyacen a la actividad locomotora, las contracciones musculares son multifacéticas, ya que pueden producir cambios en la longitud y la tensión de forma variable en el tiempo. [7] Por lo tanto, es probable que ni la longitud ni la tensión permanezcan constantes cuando el músculo está activo durante la actividad locomotora.
Contracción isométrica
Una contracción isométrica de un músculo genera tensión sin cambiar de longitud. [1] [3] [4] [5] Se puede encontrar un ejemplo cuando los músculos de la mano y el antebrazo agarran un objeto; las articulaciones de la mano no se mueven, pero los músculos generan suficiente fuerza para evitar que el objeto se caiga.
Contracción isotónica
En la contracción isotónica , la tensión en el músculo permanece constante a pesar de un cambio en la longitud del músculo. [1] [3] [4] [5] Esto ocurre cuando la fuerza de contracción de un músculo coincide con la carga total sobre el músculo.
Contracción concéntrica
En la contracción concéntrica, la tensión muscular es suficiente para superar la carga y el músculo se acorta a medida que se contrae. [8] Esto ocurre cuando la fuerza generada por el músculo excede la carga que se opone a su contracción.
Durante una contracción concéntrica, se estimula la contracción de un músculo de acuerdo con la teoría del filamento deslizante . Esto ocurre a lo largo del músculo, generando una fuerza en el origen y la inserción, lo que hace que el músculo se acorte y cambie el ángulo de la articulación. En relación con el codo , una contracción concéntrica del bíceps haría que el brazo se doblara a la altura del codo mientras la mano se movía de la pierna al hombro (un curl de bíceps ). Una contracción concéntrica del tríceps cambiaría el ángulo de la articulación en sentido contrario, enderezando el brazo y moviendo la mano hacia la pierna.
Contracción excéntrica
En la contracción excéntrica , la tensión generada mientras es isométrica es insuficiente para superar la carga externa sobre el músculo y las fibras musculares se alargan a medida que se contraen. [9] En lugar de trabajar para tirar de una articulación en la dirección de la contracción del músculo, el músculo actúa para desacelerar la articulación al final de un movimiento o controlar el reposicionamiento de una carga. Esto puede ocurrir involuntariamente (p. Ej., Cuando se intenta mover un peso demasiado pesado para que el músculo lo levante) o voluntariamente (p. Ej., Cuando el músculo está "suavizando" un movimiento o resistiendo la gravedad, como al caminar cuesta abajo). A corto plazo, el entrenamiento de fuerza con contracciones excéntricas y concéntricas parece aumentar la fuerza muscular.más que entrenar solo con contracciones concéntricas. [10] Sin embargo, el daño muscular inducido por el ejercicio también es mayor durante las contracciones de alargamiento. [11]
Durante una contracción excéntrica del músculo bíceps , el codo comienza el movimiento mientras está doblado y luego se endereza cuando la mano se aleja del hombro . Durante una contracción excéntrica del músculo tríceps , el codo inicia el movimiento recto y luego se dobla mientras la mano se mueve hacia el hombro. La desmina , la titina y otras proteínas de la línea z están implicadas en las contracciones excéntricas, pero su mecanismo se comprende poco en comparación con el ciclo de puentes cruzados en las contracciones concéntricas. [9]
Aunque el músculo está realizando una cantidad negativa de trabajo mecánico (se está trabajando en el músculo), la energía química (originalmente de oxígeno , [12] desbloqueada por la grasa o la glucosa y almacenada temporalmente en ATP ) se consume, aunque menos que se consumiría durante una contracción concéntrica de la misma fuerza. Por ejemplo, uno gasta más energía subiendo un tramo de escaleras que bajando el mismo tramo.
Los músculos que se someten a una carga excéntrica pesada sufren un mayor daño cuando se sobrecargan (como durante el desarrollo muscular o el ejercicio de entrenamiento de fuerza ) en comparación con la carga concéntrica. Cuando se utilizan contracciones excéntricas en el entrenamiento con pesas, normalmente se denominan negativas . Durante una contracción concéntrica, los miofilamentos musculares se deslizan unos sobre otros, uniendo las líneas Z. Durante una contracción excéntrica, los miofilamentosse deslizan uno al lado del otro en sentido contrario, aunque se desconoce el movimiento real de las cabezas de miosina durante una contracción excéntrica. El ejercicio con una carga excéntrica pesada puede soportar un peso mayor (los músculos son aproximadamente un 40% más fuertes durante las contracciones excéntricas que durante las concéntricas) y también resulta en un mayor daño muscular y dolor muscular de aparición tardía uno o dos días después del entrenamiento. El ejercicio que incorpora contracciones musculares excéntricas y concéntricas (es decir, que implica una fuerte contracción y una reducción controlada del peso) puede producir mayores ganancias en fuerza que las contracciones concéntricas solas. [10] [13] Si bien las contracciones excéntricas intensas no acostumbradas pueden conducir fácilmente a un sobreentrenamiento, un entrenamiento moderado puede conferir protección contra lesiones. [10]
Contracciones excéntricas en movimiento
Las contracciones excéntricas normalmente ocurren como una fuerza de frenado en oposición a una contracción concéntrica para proteger las articulaciones del daño. Durante prácticamente cualquier movimiento de rutina, las contracciones excéntricas ayudan a mantener los movimientos suaves, pero también pueden ralentizar los movimientos rápidos como un puñetazo o un lanzamiento. Parte del entrenamiento para movimientos rápidos como el lanzamiento durante el béisbol implica reducir el frenado excéntrico permitiendo que se desarrolle una mayor potencia a lo largo del movimiento.
Se están investigando las contracciones excéntricas por su capacidad para acelerar la rehabilitación de tendones débiles o lesionados. Se ha demostrado que la tendinitis de Aquiles [14] [15] y la tendinitis rotuliana [16] (también conocida como rodilla de saltador o tendonosis rotuliana) se benefician de las contracciones excéntricas de alta carga.
Vertebrados

En los animales vertebrados , hay tres tipos de tejidos musculares: esquelético, liso y cardíaco. El músculo esquelético constituye la mayor parte de la masa muscular del cuerpo y es responsable de la actividad locomotora. El músculo liso forma los vasos sanguíneos , el tracto gastrointestinal y otras áreas del cuerpo que producen contracciones sostenidas. El músculo cardíaco forma el corazón, que bombea sangre. Los músculos esqueléticos y cardíacos se denominan músculos estriados debido a su apariencia rayada bajo un microscopio, que se debe al patrón alterno altamente organizado de las bandas A y las bandas I.
Músculo esquelético
Excluyendo los reflejos, todas las contracciones de los músculos esqueléticos ocurren como resultado de un esfuerzo consciente que se origina en el cerebro . El cerebro envía señales electroquímicas a través del sistema nervioso a la neurona motora que inerva varias fibras musculares. [17] En el caso de algunos reflejos , la señal de contraerse puede originarse en la médula espinal a través de un circuito de retroalimentación con la materia gris. Otras acciones como la locomoción, la respiración y la masticación tienen un aspecto reflejo: las contracciones pueden iniciarse tanto consciente como inconscientemente.
Unión neuromuscular
Una unión neuromuscular es una sinapsis química formada por el contacto entre una neurona motora y una fibra muscular . [18] Es el sitio en el que una neurona motora transmite una señal a una fibra muscular para iniciar la contracción muscular. La secuencia de eventos que da como resultado la despolarización de la fibra muscular en la unión neuromuscular comienza cuando se inicia un potencial de acción en el cuerpo celular de una neurona motora, que luego se propaga por conducción saltatoria a lo largo de su axón hacia la unión neuromuscular. Una vez que llega al botón terminal , el potencial de acción provoca un Ca2+entrada de iones en el terminal a través de los canales de calcio dependientes de voltaje . El Ca2+El influjo hace que las vesículas sinápticas que contienen el neurotransmisor acetilcolina se fusionen con la membrana plasmática, liberando acetilcolina en la hendidura sináptica entre la terminal de la neurona motora y la unión neuromuscular de la fibra del músculo esquelético. La acetilcolina se difunde a través de la sinapsis y se une y activa los receptores nicotínicos de acetilcolina en la unión neuromuscular. La activación del receptor nicotínico abre su canal intrínseco de sodio / potasio , lo que hace que el sodio ingrese rápidamente y el potasio se filtre. Como resultado, el sarcolemainvierte la polaridad y su voltaje salta rápidamente desde el potencial de membrana en reposo de -90mV hasta un máximo de + 75mV cuando entra sodio. El potencial de membrana luego se hiperpolariza cuando sale el potasio y luego se ajusta de nuevo al potencial de membrana en reposo. Esta rápida fluctuación se denomina potencial de la placa terminal. [19] Los canales iónicos del sarcolema activados por voltaje junto a la placa terminal se abren en respuesta al potencial de la placa terminal. Son específicos de sodio y potasio y solo permiten el paso de uno. Esta onda de movimientos de iones crea el potencial de acción que se extiende desde la placa del extremo del motor en todas las direcciones. [19]Si los potenciales de acción dejan de llegar, la acetilcolina deja de liberarse del botón terminal. La acetilcolina restante en la hendidura sináptica es degradada por la acetilcolina esterasa activa o reabsorbida por el botón sináptico y no queda nada para reemplazar la acetilcolina degradada.
Acoplamiento excitación-contracción
El acoplamiento excitación-contracción es el proceso por el cual un potencial de acción muscular en la fibra muscular hace que las miofibrillas se contraigan. [20] En el músculo esquelético, el acoplamiento de excitación-contracción se basa en un acoplamiento directo entre proteínas clave, el canal de liberación de calcio del retículo sarcoplásmico (SR) (identificado como el receptor de rianodina 1 , (RYR1) y los canales de calcio de tipo L dependientes de voltaje ( identificados como receptores de dihidropiridina , DHPR). Los DHPR se encuentran en el sarcolema (que incluye el sarcolema de superficie y los túbulos transversales), mientras que los RyR residen a través de la membrana SR. La estrecha aposición de un túbulo transversal y dos regiones SR que contienen RyR se describe como una tríada y es predominantemente donde tiene lugar el acoplamiento excitación-contracción. El acoplamiento excitación-contracción ocurre cuando la despolarización de la célula del músculo esquelético da como resultado un potencial de acción muscular, que se extiende a través de la superficie celular y dentro de la red de túbulos T de la fibra muscular, despolarizando así la porción interna de la fibra muscular. La despolarización de las porciones internas activa los receptores de dihidropiridina en las cisternas terminales, que están muy cerca de los receptores de rianodina en el retículo sarcoplásmico adyacente.. Los receptores de dihidropiridina activados interactúan físicamente con los receptores de rianodina para activarlos a través de los procesos del pie (que implican cambios conformacionales que activan alostéricamente los receptores de rianodina). A medida que se abren los receptores de rianodina, Ca2+
se libera del retículo sarcoplásmico al espacio de unión local y se difunde en el citoplasma voluminoso para provocar una chispa de calcio . Tenga en cuenta que el retículo sarcoplásmico tiene una gran capacidad amortiguadora de calcio debido en parte a una proteína de unión al calcio llamada calsequestrina . La activación casi sincrónica de miles de chispas de calcio por el potencial de acción provoca un aumento de calcio en toda la célula que da lugar a la subida del transitorio de calcio . El Ca2+
liberado en el citosol se une a la troponina C por los filamentos de actina , para permitir el ciclo del puente cruzado, produciendo fuerza y, en algunas situaciones, movimiento. El sarco / retículo endoplásmico calcio-ATPasa (SERCA) bombea activamente Ca2+
de vuelta al retículo sarcoplásmico. Como Ca2+
vuelve a los niveles de reposo, la fuerza disminuye y se produce la relajación.
Teoría del filamento deslizante

La teoría del filamento deslizante describe un proceso que utilizan los músculos para contraerse. Es un ciclo de eventos repetitivos que hacen que un filamento delgado se deslice sobre un filamento grueso y genere tensión en el músculo. [21] Fue desarrollado independientemente por Andrew Huxley y Rolf Niedergerke y por Hugh Huxley y Jean Hanson en 1954. [22] [23] Fisiológicamente, esta contracción no es uniforme a través del sarcómero; la posición central de los filamentos gruesos se vuelve inestable y puede cambiar durante la contracción. Sin embargo, las acciones de proteínas elásticas como la titinase supone que mantienen una tensión uniforme a través del sarcómero y tiran del filamento grueso hacia una posición central. [24]
Ciclo de cruce de puentes
El ciclo de puentes cruzados es una secuencia de eventos moleculares que subyace a la teoría del filamento deslizante. Un puente cruzado es una proyección de miosina, que consta de dos cabezas de miosina, que se extiende desde los filamentos gruesos. [1] Cada cabeza de miosina tiene dos sitios de unión: uno para ATP y otro para actina. La unión de ATP a una cabeza de miosina separa la miosina de la actina , lo que permite que la miosina se una a otra molécula de actina. Una vez adherido, el ATP es hidrolizado por la miosina, que usa la energía liberada para moverse a la "posición amartillada", por lo que se une débilmente a una parte del sitio de unión de la actina. El resto del sitio de unión de actina está bloqueado por tropomiosina . [25]Con el ATP hidrolizado, la cabeza de miosina de tres picos contiene ahora ADP + P i . Dos Ca2+
Los iones se unen a la troponina C en los filamentos de actina. La troponina- Ca2+
El complejo hace que la tropomiosina se deslice y desbloquee el resto del sitio de unión de la actina. Desbloquear el resto de los sitios de unión de actina permite que las dos cabezas de miosina se cierren y la miosina se una fuertemente a la actina. [25] La cabeza de miosina luego libera el fosfato inorgánico e inicia un golpe de potencia, que genera una fuerza de 2 pN. El golpe de poder mueve el filamento de actina hacia adentro, acortando así el sarcómero . Luego, la miosina libera ADP pero aún permanece fuertemente unida a la actina. Al final del golpe de potencia, el ADP se libera de la cabeza de miosina, dejando la miosina unida a la actina en un estado de rigor hasta que otro ATP se une a la miosina. La falta de ATP daría como resultado el estado de rigor característico del rigor mortis. Una vez que otro ATP se une a la miosina, la cabeza de la miosina se desprenderá nuevamente de la actina y se producirá otro ciclo de puentes cruzados.
El ciclo de puentes cruzados puede continuar siempre que haya cantidades suficientes de ATP y Ca2+
en el citoplasma. [25] La terminación del ciclo del puente cruzado puede ocurrir cuando Ca2+
se bombea activamente hacia el retículo sarcoplásmico. Cuando Ca2+
ya no está presente en el filamento delgado, la tropomiosina cambia de conformación a su estado anterior para bloquear nuevamente los sitios de unión. La miosina deja de unirse al filamento delgado y el músculo se relaja. El Ca2+
Los iones abandonan la molécula de troponina para mantener el Ca2+
concentración de iones en el sarcoplasma. El bombeo activo de Ca2+
iones en el retículo sarcoplásmico crea una deficiencia en el líquido alrededor de las miofibrillas. Esto provoca la eliminación de Ca2+
iones de la troponina. Por tanto, el complejo tropomiosina-troponina vuelve a cubrir los sitios de unión de los filamentos de actina y cesa la contracción.
Graduación de las contracciones del músculo esquelético
La fuerza de las contracciones del músculo esquelético se puede dividir en general en contracciones , sumatoria y tétanos . Una contracción es un ciclo único de contracción y relajación producido por un potencial de acción dentro de la propia fibra muscular. [26] El tiempo que transcurre entre un estímulo del nervio motor y la subsiguiente contracción del músculo inervado se denomina período latente , que suele durar unos 10 ms y es causado por el tiempo que tarda en propagarse el potencial de acción del nervio, el tiempo que tarda la sustancia química transmisión en la unión neuromuscular, luego los pasos subsiguientes en el acoplamiento excitación-contracción. [27]
Si se produjera otro potencial de acción muscular antes de la relajación completa de una contracción muscular, entonces la siguiente contracción simplemente se sumará a la contracción anterior, produciendo así una suma . La suma se puede lograr de dos formas: [28] suma de frecuencia y suma de fibras múltiples . En la suma de frecuencias , la fuerza ejercida por el músculo esquelético se controla variando la frecuencia a la que los potenciales de acciónse envían a las fibras musculares. Los potenciales de acción no llegan a los músculos de forma sincrónica y, durante una contracción, alguna fracción de las fibras del músculo se activará en un momento dado. En una circunstancia típica, cuando los seres humanos están ejercitando sus músculos con tanta fuerza como son conscientemente capaces, aproximadamente un tercio de las fibras en cada uno de esos músculos se dispararán a la vez [ cita requerida ] , aunque esta proporción puede verse afectada por varios factores fisiológicos y factores psicológicos (incluidos los órganos tendinosos de Golgi y las células de Renshaw ). Este "bajo" nivel de contracción es un mecanismo de protección para prevenir la avulsión del tendón; la fuerza generada por una contracción del 95% de todas las fibras es suficiente para dañar el cuerpo. EnSuma de fibras múltiples , si el sistema nervioso central envía una señal débil para contraer un músculo, las unidades motoras más pequeñas , que son más excitables que las más grandes, se estimulan primero. A medida que aumenta la fuerza de la señal , se excitan más unidades motoras además de las más grandes, y las unidades motoras más grandes tienen hasta 50 veces la fuerza contráctil que las más pequeñas. A medida que se activan más y más unidades motoras, la fuerza de contracción muscular se vuelve progresivamente más fuerte. Un concepto conocido como el principio del tamaño permite que se produzca una gradación de la fuerza muscular durante la contracción débil en pequeños pasos, que luego se vuelven progresivamente más grandes cuando se requieren mayores cantidades de fuerza.
Finalmente, si la frecuencia de los potenciales de acción muscular aumenta de manera que la contracción muscular alcanza su fuerza máxima y se estabiliza en este nivel, entonces la contracción es un tétanos .
Relación longitud-tensión
La relación longitud-tensión relaciona la fuerza de una contracción isométrica con la longitud del músculo en el que se produce la contracción. Los músculos operan con mayor tensión activa cuando están cerca de una longitud ideal (a menudo su longitud de reposo). Cuando se estira o se acorta más allá de este (ya sea por la acción del propio músculo o por una fuerza externa), la tensión activa máxima generada disminuye. [29]Esta disminución es mínima para pequeñas desviaciones, pero la tensión cae rápidamente a medida que la longitud se desvía más del ideal. Debido a la presencia de proteínas elásticas dentro de una célula muscular (como la titina) y la matriz extracelular, a medida que el músculo se estira más allá de una longitud determinada, existe una tensión completamente pasiva, que se opone al alargamiento. Combinados, hay una fuerte resistencia a alargar un músculo activo mucho más allá del pico de tensión activa.
Relaciones fuerza-velocidad

La relación fuerza-velocidad relaciona la velocidad a la que un músculo cambia su longitud (normalmente regulada por fuerzas externas, como la carga u otros músculos) con la cantidad de fuerza que genera. La fuerza disminuye de manera hiperbólica en relación con la fuerza isométrica a medida que aumenta la velocidad de acortamiento, llegando finalmente a cero a cierta velocidad máxima. Lo contrario es válido para cuando el músculo se estira: la fuerza aumenta por encima del máximo isométrico, hasta que finalmente alcanza un máximo absoluto. Esta propiedad intrínseca del tejido muscular activo juega un papel en la amortiguación activa de las articulaciones que son accionadas por músculos opuestos simultáneamente activos. En tales casos, el perfil fuerza-velocidad mejora la fuerza producida por el músculo alargador a expensas del músculo acortador.Este favorecimiento de cualquier músculo que devuelva el equilibrio a la articulación aumenta efectivamente la amortiguación de la articulación. Además, la fuerza de la amortiguación aumenta con la fuerza muscular. De este modo, el sistema motor puede controlar activamente la amortiguación articular mediante la contracción simultánea (cocontracción) de grupos musculares opuestos.[30]
Músculo liso
Los músculos lisos se pueden dividir en dos subgrupos: de una sola unidad y de múltiples unidades . Se pueden encontrar células de músculo liso de una sola unidad en el intestino y los vasos sanguíneos. Debido a que estas células están unidas entre sí por uniones gap, pueden contraerse como un sincitio funcional . Las células de músculo liso de una sola unidad se contraen miogénicamente, lo que puede ser modulado por el sistema nervioso autónomo.
A diferencia de las células de músculo liso de una sola unidad, las células de músculo liso de múltiples unidades se encuentran en el músculo del ojo y en la base de los folículos pilosos. Las células de músculo liso de múltiples unidades se contraen al ser estimuladas por separado por nervios del sistema nervioso autónomo. Como tales, permiten un control fino y respuestas graduales, al igual que el reclutamiento de unidades motoras en el músculo esquelético.
Mecanismos de contracción del músculo liso
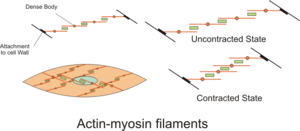
La actividad contráctil de las células del músculo liso puede ser tónica (sostenida) o fásica (transitoria) [31] y está influenciada por múltiples estímulos como la actividad eléctrica espontánea, estímulos neuronales y hormonales, cambios locales en la composición química y estiramiento. [1] Esto contrasta con la actividad contráctil de las células del músculo esquelético, que depende de una única entrada neuronal. Algunos tipos de células de músculo liso pueden generar sus propios potenciales de acción de forma espontánea, lo que suele ocurrir después de un potencial de marcapasos o de un potencial de onda lenta . Estos potenciales de acción son generados por la afluencia de Ca extracelular2+
, y no Na+
. Como los músculos esqueléticos, el Ca citosólico2+
Los iones también son necesarios para el ciclo de puentes cruzados en las células del músculo liso.
Las dos fuentes de Ca citosólico2+
en las células del músculo liso son el Ca extracelular2+
entrando a través de los canales de calcio y el Ca2+
iones que se liberan del retículo sarcoplásmico. La elevación del Ca citosólico2+
da como resultado más Ca2+
unión a calmodulina , que luego se une y activa la quinasa de cadena ligera de miosina . El complejo calcio-calmodulina-miosina-quinasa de cadena ligera fosforila la miosina en las cadenas ligeras de miosina de 20 kilodalton (kDa) en el residuo de aminoácido-serina 19, iniciando la contracción y activando la miosina ATPasa . A diferencia de las células del músculo esquelético, las células del músculo liso carecen de troponina, a pesar de que contienen la proteína de filamento delgado tropomiosina y otras proteínas notables: caldesmon y calponina. Por lo tanto, las contracciones del músculo liso son iniciadas por el Ca2+
fosforilación activada de miosina en lugar de Ca2+
unión al complejo de troponina que regula los sitios de unión de miosina en actina como en los músculos esqueléticos y cardíacos.
La terminación del ciclo del puente cruzado (y dejar el músculo en estado de enganche) ocurre cuando la fosfatasa de cadena ligera de miosina elimina los grupos fosfato de las cabezas de miosina. La fosforilación de las cadenas ligeras de miosina de 20 kDa se correlaciona bien con la velocidad de acortamiento del músculo liso. Durante este período, hay una rápida explosión de utilización de energía medida por el consumo de oxígeno. A los pocos minutos del inicio, el nivel de calcio disminuye marcadamente, la fosforilación de las cadenas ligeras de miosina de 20 kDa disminuye y la utilización de energía disminuye; sin embargo, se mantiene la fuerza en el músculo liso tónico. Durante la contracción del músculo, se forman puentes cruzados de ciclo rápido entre la actina activada y la miosina fosforilada, lo que genera fuerza. Se plantea la hipótesis de que el mantenimiento de la fuerza resulta de "puentes de cierre" desfosforilados.que giran lentamente y mantienen la fuerza. Varias quinasas comoSe cree que la rho quinasa , DAPK3 y la proteína quinasa C participan en la fase sostenida de contracción, y Ca2+
el flujo puede ser significativo.
Neuromodulación
Aunque las contracciones del músculo liso son miogénicas, el sistema nervioso autónomo puede modular la velocidad y la fuerza de sus contracciones . Las fibras nerviosas posganglionares del sistema nervioso parasimpático liberan el neurotransmisor acetilcolina, que se une a los receptores muscarínicos de acetilcolina (mAChR) en las células del músculo liso. Estos receptores son receptores metabotrópicos o acoplados a proteína G que inician una cascada de segundos mensajeros. Por el contrario, las fibras nerviosas posganglionares del sistema nervioso simpáticoliberan los neurotransmisores epinefrina y norepinefrina, que se unen a receptores adrenérgicos que también son metabotrópicos. Los efectos exactos sobre el músculo liso dependen de las características específicas del receptor activado; tanto la entrada parasimpática como la simpática pueden ser excitadoras (contráctiles) o inhibidoras (relajantes).
Músculo cardíaco

Hay dos tipos de células del músculo cardíaco: autorrítmicas y contráctiles. Las células autorrítmicas no se contraen, sino que marcan el ritmo de contracción de otras células del músculo cardíaco, que pueden ser moduladas por el sistema nervioso autónomo. Por el contrario, las células de los músculos contráctiles (cardiomiocitos) constituyen la mayor parte del músculo cardíaco y pueden contraerse.
Acoplamiento excitación-contracción
Tanto en el acoplamiento de excitación-contracción (CE) del músculo esquelético como en el cardíaco , se producen procesos de conducción de despolarización y liberación de Ca 2+ . Sin embargo, aunque las proteínas involucradas son similares, son distintas en estructura y regulación. Los receptores de dihidropiridina (DHPR) están codificados por diferentes genes y los receptores de rianodina (RyR) son isoformas distintas. Además, el DHPR entra en contacto con RyR1 (isoforma principal de RyR en el músculo esquelético) para regular la liberación de Ca 2+ en el músculo esquelético, mientras que el canal de calcio tipo L (DHPR en los miocitos cardíacos) y RyR2 (isoforma principal de RyR en el músculo cardíaco) no lo son físicamente. acoplados en el músculo cardíaco, pero enfrentados entre sí por un acoplamiento de unión.[32]
A diferencia del músculo esquelético, se cree que el acoplamiento de CE en el músculo cardíaco depende principalmente de un mecanismo llamado liberación de calcio inducida por calcio , [33] que se basa en la estructura de unión entre el túbulo T y el retículo sarcoplásmico. Junctophilin-2 (JPH2) es esencial para mantener esta estructura, así como la integridad de T-túbulo . [34] [35] [36] Otra proteína, la proteína accesoria del receptor 5 (REEP5), funciona para mantener la morfología normal de la RS de unión. [37]Los defectos del acoplamiento de la unión pueden resultar de deficiencias de cualquiera de las dos proteínas. Durante el proceso de liberación de calcio inducida por calcio, los RyR2 son activados por un disparador de calcio, que es provocado por el flujo de Ca 2+ a través de los canales de calcio de tipo L. Después de esto, el músculo cardíaco tiende a exhibir estructuras diadiales , en lugar de tríadas .
El acoplamiento excitación-contracción en las células del músculo cardíaco se produce cuando las células marcapasos en el nódulo sinoauricular o el nódulo auriculoventricular inician un potencial de acción y se conduce a todas las células del corazón a través de uniones gap . El potencial de acción viaja a lo largo de la membrana superficial hacia los túbulos T (estos últimos no se observan en todos los tipos de células cardíacas) y la despolarización causa Ca extracelular.2+
para ingresar a la célula a través de los canales de calcio de tipo L y posiblemente el intercambiador de sodio-calcio (NCX) durante la primera parte de la fase de meseta . Aunque este influjo de Ca 2+ solo cuenta para aproximadamente el 10% del Ca 2+ necesario para la activación, es relativamente mayor que el del músculo esquelético. Este Ca2+
El influjo provoca un pequeño aumento local en el Ca intracelular.2+
. El aumento de Ca intracelular2+
es detectado por RyR2 en la membrana del retículo sarcoplásmico, que libera Ca2+
en una respuesta fisiológica de retroalimentación positiva . Esta retroalimentación positiva se conoce como liberación de calcio inducida por calcio [33] y da lugar a chispas de calcio ( Ca2+
chispas [38] ). La suma espacial y temporal de ~ 30.000 Ca2+
las chispas dan un aumento en toda la célula de la concentración de calcio citoplásmico. [39] El aumento de calcio citosólico que sigue al flujo de calcio a través de la membrana celular y el retículo sarcoplásmico es moderado por tampones de calcio , que se unen a una gran proporción de calcio intracelular. Como resultado, un gran aumento en el calcio total conduce a un aumento relativamente pequeño en el Ca libre.2+
. [40]
El calcio citoplasmático se une a la troponina C, moviendo el complejo de tropomiosina fuera del sitio de unión de la actina, lo que permite que la cabeza de la miosina se una al filamento de actina. A partir de este momento, el mecanismo contráctil es esencialmente el mismo que para el músculo esquelético (arriba). Brevemente, usando hidrólisis de ATP, la cabeza de miosina tira del filamento de actina hacia el centro del sarcómero.

Después de la sístole, el calcio intracelular es absorbido por la bomba de ATPasa del retículo sarco / endoplásmico (SERCA) de regreso al retículo sarcoplásmico listo para que comience el siguiente ciclo. El calcio también es expulsado de la célula principalmente por el intercambiador de sodio-calcio (NCX) y, en menor grado, una ATPasa de calcio de la membrana plasmática . Las mitocondrias también absorben algo de calcio. [41] Una enzima, fosfolamban , sirve como freno para SERCA. A frecuencias cardíacas bajas, el fosfolambán está activo y ralentiza la actividad de la ATPasa para que el Ca2+
no tiene que salir de la celda por completo. A frecuencias cardíacas elevadas, el fosfolambán se fosforila y se desactiva, por lo que consume la mayor parte del Ca2+
del citoplasma al retículo sarcoplásmico. Una vez más, los tampones de calcio moderan esta caída en Ca2+
concentración, lo que permite una disminución relativamente pequeña de Ca libre2+
concentración en respuesta a un gran cambio en el calcio total. La caída de Ca2+
La concentración permite que el complejo de troponina se disocie del filamento de actina, poniendo fin a la contracción. El corazón se relaja, permitiendo que los ventrículos se llenen de sangre y comiencen de nuevo el ciclo cardíaco.
Invertebrados
Músculos circulares y longitudinales
En anélidos como las lombrices de tierra y las sanguijuelas , las células de los músculos circulares y longitudinales forman la pared corporal de estos animales y son responsables de su movimiento. [42] En una lombriz de tierra que se mueve a través de un suelo, por ejemplo, las contracciones de los músculos circulares y longitudinales ocurren recíprocamente mientras que el fluido celómico sirve como un hidroesqueleto al mantener la turgencia de la lombriz de tierra. [43]Cuando los músculos circulares de los segmentos anteriores se contraen, la parte anterior del cuerpo del animal comienza a contraerse radialmente, lo que empuja el fluido celómico incompresible hacia adelante y aumenta la longitud del animal. Como resultado, la parte delantera del animal se mueve hacia adelante. A medida que el extremo delantero de la lombriz de tierra se ancla y los músculos circulares de los segmentos anteriores se relajan, una ola de contracciones musculares longitudinales pasa hacia atrás, lo que empuja hacia adelante el resto del cuerpo del animal. [42] [43] Estas ondas alternas de contracciones circulares y longitudinales se denominan peristalsis , que subyace al movimiento progresivo de las lombrices de tierra.
Músculos oblicuamente estriados
Los invertebrados, como anélidos, moluscos y nematodos , poseen músculos estriados oblicuamente, que contienen bandas de filamentos gruesos y delgados que están dispuestos helicoidalmente en lugar de transversalmente, como en los músculos esqueléticos o cardíacos de los vertebrados. [44] En los bivalvos , los músculos estriados oblicuamente pueden mantener la tensión durante períodos prolongados sin utilizar demasiada energía. Los bivalvos usan estos músculos para mantener sus caparazones cerrados.
Músculos asincrónicos

Los insectos avanzados como avispas , moscas , abejas y escarabajos poseen músculos asincrónicos que constituyen los músculos de vuelo en estos animales. [44] Estos músculos de vuelo a menudo se denominan músculos fibrilares porque contienen miofibrillas gruesas y llamativas. [45] Una característica notable de estos músculos es que no requieren estimulación para cada contracción muscular. De ahí que se denominen músculos asincrónicos.porque el número de contracciones en estos músculos no se corresponde (o sincroniza) con el número de potenciales de acción. Por ejemplo, el músculo del ala de una mosca atada puede recibir potenciales de acción a una frecuencia de 3 Hz, pero puede latir a una frecuencia de 120 Hz. [44] El latido de alta frecuencia es posible porque los músculos están conectados a un sistema resonante , que es impulsado a una frecuencia natural de vibración.
Historia
En 1780, Luigi Galvani descubrió que los músculos de las ancas de las ranas muertas se contraían cuando eran golpeados por una chispa eléctrica. [47] Esta fue una de las primeras incursiones en el estudio de la bioelectricidad , un campo que aún estudia los patrones eléctricos y las señales en tejidos como nervios y músculos.
En 1952, se acuñó el término acoplamiento excitación-contracción para describir el proceso fisiológico de convertir un estímulo eléctrico en una respuesta mecánica. [20] Este proceso es fundamental para la fisiología muscular, por lo que el estímulo eléctrico suele ser un potencial de acción y la respuesta mecánica es la contracción. El acoplamiento excitación-contracción puede estar desregulado en muchas enfermedades. Aunque el acoplamiento excitación-contracción se conoce desde hace más de medio siglo, sigue siendo un área activa de investigación biomédica. El esquema general es que llega un potencial de acción para despolarizar la membrana celular. Por mecanismos específicos del tipo de músculo, esta despolarización da como resultado un aumento del calcio citosólico.eso se llama calcio transitorio. Este aumento de calcio activa las proteínas contráctiles sensibles al calcio que luego usan ATP para provocar el acortamiento celular.
El mecanismo de la contracción muscular eludió a los científicos durante años y requiere investigación y actualización continuas. [48] La teoría del filamento deslizante fue desarrollada independientemente por Andrew F. Huxley y Rolf Niedergerke y por Hugh Huxley y Jean Hanson . Sus hallazgos fueron publicados como dos artículos consecutivos publicados en la edición del 22 de mayo de 1954 de Nature bajo el tema común "Cambios estructurales en el músculo durante la contracción". [22] [23]
Ver también
- Términos anatómicos de movimiento
- liberación de calcio inducida por calcio
- Potencial de acción cardíaco
- Calambre
- Distonía
- Fisiología del Ejercicio
- Fasciculación
- Modelo muscular de Hill
- Idiota hipnico
- Pruebas musculares in vitro
- La paradoja de Lombard
- Mioclonías
- Rigor mortis
- Espasmo
- Contraccion uterina
Referencias
- ^ a b c d e f g h i j k l m n o Widmaier, Eric P .; Raff, Hersel; Strang, Kevin T. (2010). "Músculo". Fisiología humana de Vander: los mecanismos de la función corporal (12ª ed.). Nueva York, NY: McGraw-Hill. págs. 250-291. ISBN 978-0-321-98122-6.
- ↑ Silverthorn, Dee Unglaub (2016). "Músculos". Fisiología humana: un enfoque integrado (7ª ed.). San Francisco, CA: Pearson. págs. 377–416. ISBN 978-0-321-98122-6.
- ↑ a b c d e f Aidley, David J. (1998). "Mecánica y energética de la contracción muscular". La fisiología de las células excitables (4ª ed.). Nueva York, NY: Cambridge University Press. págs. 323–335 . ISBN 978-0-521-57421-1.
- ↑ a b c d e f Sircar, Sabyasachi (2008). "Elasticidad muscular". Principios de fisiología médica (1ª ed.). Nueva York, NY: Thieme. pag. 113. ISBN 978-1-588-90572-7.
- ^ a b c d e f Bullock, John; Boyle, Joseph; Wang, Michael B. (2001). "Contracción muscular". Fisiología NMS . 578 (4ª ed.). Baltimore, Maryland: Lippincott Williams y Wilkins. págs. 37–56.
- ↑ a b Kumar, Shrawan (2008). "Introducción y terminología". En Shrawan Kumar (ed.). Fuerza muscular (1ª ed.). Boca Raton, FL: CRC Press. pag. 113. ISBN 978-0-415-36953-4.
- ↑ a b Biewener, Andrew A. (2003). "Músculos y esqueletos: los componentes básicos del movimiento animal". Locomoción animal . Serie de Biología Animal de Oxford. Nueva York, NY: Oxford University Press. págs. 15–45. ISBN 978-0-198-50022-3.
- ^ Faulkner JA (2003). "Terminología para las contracciones de los músculos durante el acortamiento, mientras que isométrica y durante el alargamiento". Revista de fisiología aplicada . 95 (2): 455–459. doi : 10.1152 / japplphysiol.00280.2003 . PMID 12851415 .
- ^ a b "Tipos de contracciones" . 2006-05-31 . Consultado el 2 de octubre de 2007 .
- ↑ a b c Colliander EB, Tesch PA (1990). "Efectos de las acciones musculares excéntricas y concéntricas en el entrenamiento de resistencia". Acta Physiol. Scand . 140 (1): 31–9. doi : 10.1111 / j.1748-1716.1990.tb08973.x . PMID 2275403 .
- ^ Nikolaidis MG, Kyparos A, Spanou C, Paschalis V, Theodorou AA, Vrabas IS (2012). "Biología redox del ejercicio: una consideración integradora y comparativa de algunos problemas pasados por alto" . J. Exp. Biol . 215 (Pt 10): 1615-25. doi : 10.1242 / jeb.067470 . PMID 22539728 .
- ^ Schmidt-Rohr, K. (2020). "El oxígeno es el de alta energía Encendido molécula compleja multicelular Vida: Las correcciones fundamentales a la tradicional Bioenergética ACS Omega 5 :. 2221-2233 http://dx.doi.org/10.1021/acsomega.9b03352
- ^ Brooks, GA; Fahey, TD; White, TP (1996). Fisiología del ejercicio: bioenergética humana y sus aplicaciones. (2ª ed.) . Mayfield Publishing Co.
- ^ Alfredson, H; Pietilä, T; Jonsson, P; Lorentzon, R (1998). "Entrenamiento excéntrico de los músculos de la pantorrilla con carga pesada para el tratamiento de la tendinosis de Aquiles crónica" (PDF) . The American Journal of Sports Medicine . 26 (3): 360–6. doi : 10.1177 / 03635465980260030301 . PMID 9617396 . S2CID 30259362 .
- ^ Satyendra L, Byl N (2006). "Efectividad de la fisioterapia para la tendinopatía de Aquiles: una revisión basada en la evidencia de ejercicios excéntricos" . Isocinética y ciencia del ejercicio . 14 (1): 71–80. doi : 10.3233 / IES-2006-0223 .
- ^ Cannell LJ, Taunton JE, Clement DB, Smith C, Khan KM (2001). "Un ensayo clínico aleatorizado de la eficacia de las sentadillas o los ejercicios de extensión / curl de piernas para tratar la rodilla de saltador diagnosticada clínicamente en atletas: estudio piloto" . Br J Sports Med . 35 (1): 60–4. doi : 10.1136 / bjsm.35.1.60 . PMC 1724276 . PMID 11157465 .
- ^ Tassinary; Cacioppo (2000). "El sistema Skeletomotor: electromiografía de superficie". En Cacioppo, John T .; Tassinary, Luois G .; Berntson, Gary G. (eds.). Manual de psicofisiología (Segunda ed.). Cambridge: Cambridge University Press. ISBN 978-0-521-62634-7.
- ^ Levitan, Irwin; Kaczmarek, Leonard (19 de agosto de 2015). "Comunicación intercelular". La neurona: biología celular y molecular (4ª ed.). Nueva York, NY: Oxford University Press. págs. 153–328. ISBN 978-0199773893.
- ↑ a b Saladin, Kenneth S., Stephen J. Sullivan y Christina A. Gan. Anatomía y fisiología: la unidad de forma y función. 7ª ed. Nueva York: McGraw-Hill Education, 2015. Imprimir.
- ↑ a b Sandow A (1952). "Acoplamiento excitación-contracción en la respuesta muscular" . Yale J Biol Med . 25 (3): 176–201. PMC 2599245 . PMID 13015950 .
- ^ Saladino, Kenneth (2012). Anatomía y fisiología: la unidad de forma y función . Nueva York: McGraw Hill. ISBN 978-0-07-337825-1.
- ↑ a b Huxley AF, Niedergerke R (1954). "Cambios estructurales en el músculo durante la contracción: microscopía de interferencia de fibras musculares vivas". Naturaleza . 173 (4412): 971–973. Código bibliográfico : 1954Natur.173..971H . doi : 10.1038 / 173971a0 . PMID 13165697 . S2CID 4275495 .
- ↑ a b Huxley H, Hanson J (1954). "Cambios en las estrías transversales del músculo durante la contracción y el estiramiento y su interpretación estructural". Naturaleza . 173 (4412): 973–976. Código Bibliográfico : 1954Natur.173..973H . doi : 10.1038 / 173973a0 . PMID 13165698 . S2CID 4180166 .
- ^ Horowits R, Podolsky RJ (noviembre de 1987). "La estabilidad posicional de los filamentos gruesos en el músculo esquelético activado depende de la longitud del sarcómero: evidencia del papel de los filamentos de titina" . J. Cell Biol . 105 (5): 2217–23. doi : 10.1083 / jcb.105.5.2217 . PMC 2114850 . PMID 3680378 .
- ^ a b c Enoka, Roger M .; Pearson, Keir G. (2013). "La unidad motora y la acción muscular". En Eric R. Kandel; James H. Schwartz; Thomas M. Jessell; Steven A. Siegelbaum; AJ Hudspeth (eds.). Principios de la ciencia neuronal (5ª ed.). Nueva York, NY: McGraw-Hill Medical. págs. 768–789. ISBN 978-0-071-39011-8.
- ^ Feher, Joseph (2012). "Capítulo 3.4: Mecánica del músculo esquelético". Fisiología humana cuantitativa: una introducción . Serie de Prensa Académica en Ingeniería Biomédica (1ª ed.). Nueva York, NY: Academic Press. págs. 239–248. ISBN 978-0-123-82163-8.
- ^ Khurana, Indu (2006). "Características de la excitabilidad y contractilidad muscular". Libro de texto de fisiología médica (1ª ed.). Elsevier. págs. 101–2.
- ^ Shwedyk, E .; Balasubramanian, R .; Scott, Enfermera registrada (1977). "Un modelo no estacionario para el electromiograma". Transacciones IEEE sobre Ingeniería Biomédica . 24 (5): 417–424. doi : 10.1109 / TBME.1977.326175 . PMID 892834 . S2CID 1770255 .
- ^ Gordon AM, Huxley AF, Julian FJ (1966). "La variación en la tensión isométrica con la longitud del sarcómero en las fibras musculares de los vertebrados" . J. Physiol . 184 (1): 170–92. doi : 10.1113 / jphysiol.1966.sp007909 . PMC 1357553 . PMID 5921536 .
- ^ Heitmann, Stewart; Helechos, norma; Breakpsear, Michael (2011). "La cocontracción muscular modula la amortiguación y la estabilidad articular en una extremidad biomecánica de tres enlaces" . Fronteras en Neurorobótica . 5 : 5. doi : 10.3389 / fnbot.2011.00005 . ISSN 1662-5218 . PMC 3257849 . PMID 22275897 .
- ^ Zhang, Y; Hermanson, ME; Eddinger, TJ (2013). "La contracción tónica y fásica del músculo liso no está regulada por la vía PKCα - CPI-17 en el antro y el fondo de estómago de los cerdos" . PLOS ONE . 8 (9): e74608. doi : 10.1371 / journal.pone.0074608 . PMC 3776813 . PMID 24058600 .
- ↑ Martonosi, Anthony N .; Pikula, Slawomir (2003). "La red de regulación del calcio en el músculo" . Acta Biochimica Polonica . 50 (1): 1–30. doi : 10.18388 / abp.2003_3711 . ISSN 0001-527X . PMID 12673344 .
- ↑ a b Fabiato, A. (1983). "Liberación de calcio inducida por calcio del retículo sarcoplásmico cardíaco". Revista estadounidense de fisiología . 245 (1): C1-14. doi : 10.1152 / ajpcell.1983.245.1.C1 . PMID 6346892 .
- ^ Guo, Ang; Zhang, Xiaoying; Iyer, Venkat Ramesh; Chen, Biyi; Zhang, Caimei; Kutschke, William J .; Weiss, Robert M .; Franzini-Armstrong, Clara; Song, Long-Sheng (19 de agosto de 2014). "La sobreexpresión de junctophilin-2 no mejora la función basal, pero atenúa el desarrollo de insuficiencia cardíaca después de estrés cardíaco" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 111 (33): 12240-12245. doi : 10.1073 / pnas.1412729111 . ISSN 1091-6490 . PMC 4143026 . PMID 25092313 .
- ^ Wei, Sheng; Guo, Ang; Chen, Biyi; Kutschke, William; Xie, Yu-Ping; Zimmerman, Kathy; Weiss, Robert M .; Anderson, Mark E .; Cheng, Heping; Song, Long-Sheng (20 de agosto de 2010). "Remodelación del túbulo T durante la transición de hipertrofia a insuficiencia cardíaca" . Investigación de circulación . 107 (4): 520–531. doi : 10.1161 / CIRCRESAHA.109.212324 . ISSN 1524-4571 . PMC 2927862 . PMID 20576937 .
- ^ Takeshima, H .; Komazaki, S .; Nishi, M .; Iino, M .; Kangawa, K. (julio de 2000). "Junctofilinas: una nueva familia de proteínas del complejo de la membrana de unión" . Célula molecular . 6 (1): 11-22. doi : 10.1016 / s1097-2765 (00) 00003-4 . ISSN 1097-2765 . PMID 10949023 .
- ^ Yao, Lei; Xie, Duanyang; Geng, Li; Shi, Dan; Huang, Jian; Wu, Yufei; Lv, Fei; Liang, Dandan; Li, Li; Liu, Yi; Li, junio (3 de febrero de 2018). "REEP5 (proteína accesoria del receptor 5) actúa como un escultor de membrana de retículo sarcoplásmico para modular la función cardíaca" . Revista de la Asociación Estadounidense del Corazón . 7 (3). doi : 10.1161 / JAHA.117.007205 . ISSN 2047-9980 . PMC 5850239 . PMID 29431104 .
- ^ Cheng H, Lederer WJ, Cannell MB (octubre de 1993). "Chispas de calcio: eventos elementales subyacentes al acoplamiento de excitación-contracción en el músculo cardíaco". Ciencia . 262 (5134): 740–4. Código Bibliográfico : 1993Sci ... 262..740C . doi : 10.1126 / science.8235594 . PMID 8235594 .
- ^ Cannell MB, Cheng H, Lederer WJ (noviembre de 1994). "No uniformidades espaciales en Ca2+i durante la excitación-contracción de acoplamiento en los miocitos cardíacos" . Biophys J. . 67 (5):. 1942-1956 bibcode : 1994BpJ .... 67.1942C . doi : 10.1016 / S0006-3495 (94) 80677-0 . PMC 1225569 . PMID 7858131 .
- ^ M., Bers, D. (2001). Acoplamiento excitación-contracción y fuerza contráctil cardíaca (2ª ed.). Dordrecht: Kluwer Academic Publishers. ISBN 9780792371571. OCLC 47659382 .
- ^ Crespo LM, Grantham CJ, Cannell MB (junio de 1990). "Cinética, estequiometría y papel del mecanismo de intercambio de Na-Ca en miocitos cardíacos aislados". Naturaleza . 345 (6276): 618–21. Código Bibliográfico : 1990Natur.345..618C . doi : 10.1038 / 345618a0 . PMID 2348872 . S2CID 4348240 .
- ↑ a b Hillis, David M .; Sadava, David E .; Precio, Mary V. (2014). "Músculo y movimiento". Principios de vida (2ª ed.). Sunderland, MA: Sinauer Associates. págs. 681–698. ISBN 978-1-464-10947-8.
- ↑ a b Gardner, CR (1976). "El control neuronal de la locomoción en la lombriz de tierra". Reseñas biológicas de la Sociedad Filosófica de Cambridge . 51 (1): 25–52. doi : 10.1111 / j.1469-185X.1976.tb01119.x . PMID 766843 . S2CID 9983649 .
- ↑ a b c Alexander, R. McNeill (2003). "Músculo, el motor". Principios de locomoción animal (2ª ed.). Princeton, Nueva Jersey: Princeton University Press. págs. 15–37. ISBN 978-0-691-12634-0.
- ^ Josephson, RK; Malamud, JG; Stokes, RD (15 de septiembre de 2000). "Músculo asincrónico: una cartilla" . Revista de Biología Experimental . 203 (18): 2713–2722. doi : 10.1242 / jeb.203.18.2713 . ISSN 0022-0949 . PMID 10952872 .
- ^ David Ames Wells, La ciencia de las cosas comunes: una explicación familiar de la primera , 323 páginas ( página 290 )
- ^ Whittaker, ET (1951), Una historia de las teorías del éter y la electricidad. Vol 1 , Nelson, Londres
- ^ Huxley, HE (abril de 2000). "Experimentos pasados, presentes y futuros sobre el músculo" . Transacciones filosóficas: ciencias biológicas . 355 (1396): 539–543. doi : 10.1098 / rstb.2000.0595 . JSTOR 3066716 . PMC 1692762 . PMID 10836507 .
Lectura adicional
- Saladin, Kenneth S., Stephen J. Sullivan y Christina A. Gan. (2015). Anatomía y fisiología: la unidad de forma y función. 7ª ed. Nueva York: McGraw-Hill Education.
- Krans, JL (2010) La teoría del filamento deslizante de la contracción muscular. Educación de la naturaleza 3 (9): 66
Enlaces externos
- Modelo de filamento deslizante de contracción muscular
- Animación: contracción del miofilamento