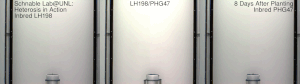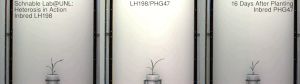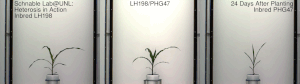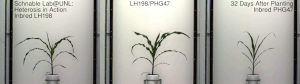
La heterosis , el vigor híbrido o la mejora de la exogamia es la función mejorada o aumentada de cualquier cualidad biológica en una descendencia híbrida . Una descendencia es heterótica si sus rasgos se mejoran como resultado de mezclar las contribuciones genéticas de sus padres. Estos efectos pueden deberse a la herencia mendeliana o no mendeliana .
Definiciones
Al proponer el término heterosis para reemplazar el antiguo término heterocigosis , GH Shull intentó evitar limitar el término a los efectos que pueden explicarse por la heterocigosidad en la herencia mendeliana. [1]
El vigor fisiológico de un organismo, manifestado en su rapidez de crecimiento, su altura y robustez general, se correlaciona positivamente con el grado de disimilitud en los gametos por cuya unión se formó el organismo ... Cuanto más numerosas sean las diferencias entre los gametos que se unen - en menos dentro de ciertos límites - mayor en general es la cantidad de estimulación ... Estas diferencias no necesitan ser mendelianas en su herencia ... Para evitar la implicación de que todas las diferencias genotípicas que estimulan la división celular, el crecimiento y otras actividades fisiológicas de un organismo son Mendeliano en su herencia y también para ganar brevedad en la expresión sugiero ... que se adopte la palabra 'heterosis'.
La heterosis a menudo se discute como lo opuesto a la depresión endogámica , aunque las diferencias en estos dos conceptos se pueden ver en consideraciones evolutivas como el papel de la variación genética o los efectos de la deriva genética en poblaciones pequeñas sobre estos conceptos. La depresión por consanguinidad ocurre cuando los padres emparentados tienen hijos con rasgos que influyen negativamente en su aptitud en gran parte debido a la homocigosidad . En tales casos, el cruzamiento externo debería resultar en heterosis.
No todos los cruces resultan en heterosis. Por ejemplo, cuando un híbrido hereda rasgos de sus padres que no son totalmente compatibles, la aptitud puede verse reducida. Esta es una forma de depresión exógena .
Dominio versus sobredominio
Dominio versus sobredominio es una controversia científica en el campo de la genética que ha persistido durante más de un siglo. [2] Estas dos hipótesis alternativas se establecieron por primera vez en 1908.
Base genética
Cuando una población es pequeña o consanguínea, tiende a perder diversidad genética. La depresión por endogamia es la pérdida de aptitud debido a la pérdida de diversidad genética. Las cepas endogámicas tienden a ser homocigotas para los alelos recesivos que son levemente dañinos (o producen un rasgo que no es deseable desde el punto de vista del criador). La heterosis o vigor híbrido, por otro lado, es la tendencia de las cepas exógamas a superar a ambos padres consanguíneos en aptitud.
La cría selectiva de plantas y animales, incluida la hibridación, comenzó mucho antes de que se comprendieran los principios científicos subyacentes. A principios del siglo XX, después de que se entendieran y aceptaran las leyes de Mendel , los genetistas se comprometieron a explicar el vigor superior de muchos híbridos de plantas. Se desarrollaron dos hipótesis en competencia, que no son mutuamente excluyentes: [3]
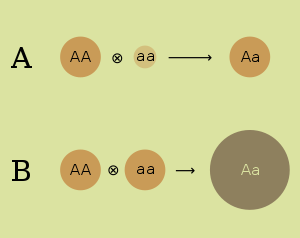
- Hipótesis de dominancia . La hipótesis de dominancia atribuye la superioridad de los híbridos a la supresión de alelos recesivos indeseables de uno de los padres por alelos dominantes del otro. Atribuye el bajo rendimiento de las cepas consanguíneas a la pérdida de diversidad genética, y las cepas se vuelven puramente homocigotas en muchos loci. La hipótesis de la dominancia fue expresada por primera vez en 1908 por el genetista Charles Davenport . [4] Bajo la hipótesis de dominancia, se espera que los alelos deletéreos se mantengan en una población de apareamiento aleatorio en un equilibrio de selección-mutación que dependería de la tasa de mutación, el efecto de los alelos y el grado en que los alelos se expresan en heterocigotos. [5]
- Hipótesis de sobredominio . Ciertas combinaciones de alelos que se pueden obtener cruzando dos cepas consanguíneas son ventajosas en el heterocigoto . La hipótesis de sobredominio atribuye la ventaja de los heterocigotos a la supervivencia de muchos alelos que son recesivos y dañinos en los homocigotos. Atribuye el bajo rendimiento de las cepas consanguíneas a un alto porcentaje de estas dañinas recesivas. La hipótesis de sobredominio fue desarrollada de forma independiente por Edward M. East (1908) [6] y George Shull (1908). [7]Se espera que la variación genética en un locus sobredominante se mantenga equilibrando la selección. La alta idoneidad de los genotipos heterocigotos favorece la persistencia de un polimorfismo alélico en la población. [5]
La dominancia y la sobredominancia tienen diferentes consecuencias para el perfil de expresión génica de los individuos. Si el sobredominio es la causa principal de las ventajas de aptitud física de la heterosis, entonces debería haber una sobreexpresión de ciertos genes en la descendencia heterocigótica en comparación con los padres homocigotos. Por otro lado, si la dominancia es la causa, menos genes deberían estar subexpresados en la descendencia heterocigota en comparación con los padres. Además, para cualquier gen dado, la expresión debería ser comparable a la observada en el más apto de los dos padres.
Retrospectiva histórica
El genetista de poblaciones James Crow (1916–2012) creía, en su juventud, que la sobredominancia era un factor importante para el vigor híbrido. En 1998 publicó una revisión retrospectiva de la ciencia en desarrollo. [8] Según Crow, la demostración de varios casos de ventaja heterocigotaen Drosophila y otros organismos primero causó un gran entusiasmo por la teoría de la sobredominancia entre los científicos que estudian la hibridación de plantas. Pero el sobredominio implica que los rendimientos en una cepa consanguínea deberían disminuir a medida que se seleccionan las cepas consanguíneas para el rendimiento de sus cruces híbridos, a medida que aumenta la proporción de recesivos dañinos en la población consanguínea. A lo largo de los años, la experimentación en genética vegetal ha demostrado que ocurre lo contrario, que los rendimientos aumentan tanto en las cepas consanguíneas como en los híbridos, lo que sugiere que la dominancia por sí sola puede ser adecuada para explicar el rendimiento superior de los híbridos. Solo se han informado unos pocos casos concluyentes de sobredominio en toda la genética. Desde la década de 1980, a medida que aumenta la evidencia experimental, la teoría de la dominancia ha vuelto.
Crow escribió:
La opinión actual ... es que la hipótesis de dominancia es la principal explicación de la disminución de la consanguinidad y [del] alto rendimiento de los híbridos. Hay poca evidencia estadística de las contribuciones del sobredominio y la epistasis . Pero si los mejores híbridos están recibiendo un impulso adicional de la sobredominancia o contribuciones epistáticas favorables sigue siendo una pregunta abierta. [8]
Controversia
El término heterosis a menudo causa confusión e incluso controversia, particularmente en la cría selectiva de animales domésticos , porque a veces se afirma (incorrectamente) que todas las plantas y animales cruzados son "genéticamente superiores" a sus padres, debido a la heterosis, [ cita requerida ] . pero existen dos problemas con esta afirmación:
- Primero, según un artículo publicado en la revista Genome Biology , " superioridad genética " es un término mal definido y no es una terminología generalmente aceptada dentro del campo científico de la genética. [9] Un término relacionado con la aptitud está bien definido, pero rara vez se puede medir directamente. En cambio, los científicos usan cantidades objetivas y medibles, como la cantidad de semillas que produce una planta, la tasa de germinación de una semilla o el porcentaje de organismos que sobreviven hasta la edad reproductiva. [10] Desde esta perspectiva, mestizalas plantas y los animales que presentan heterosis pueden tener rasgos "superiores", pero esto no necesariamente equivale a ninguna evidencia de absoluta "superioridad genética". El uso del término "superioridad" es común, por ejemplo, en el mejoramiento de cultivos, donde se entiende bien que significa una planta más robusta y de mejor rendimiento para la agricultura. Tal planta puede rendir mejor en una granja, pero probablemente tenga dificultades para sobrevivir en la naturaleza, lo que hace que este uso sea susceptible de interpretaciones erróneas. En genética humana, cualquier cuestión de "superioridad genética" es aún más problemática debido a las implicaciones históricas y políticas de tal afirmación. Algunos incluso pueden llegar a describirlo como un juicio de valor cuestionable en el ámbito de la política, no de la ciencia. [9]
- En segundo lugar, no todos los híbridos presentan heterosis (ver depresión por exogamia ).
Un ejemplo de los juicios de valor ambiguos impuestos a los híbridos y al vigor híbrido es la mula . Si bien las mulas casi siempre son infértiles, se las valora por una combinación de resistencia y temperamento que es diferente a la de sus padres, el caballo o el burro. Si bien estas cualidades pueden hacerlos "superiores" para usos particulares de los humanos, el problema de la infertilidad implica que estos animales probablemente se extinguirían sin la intervención de los humanos a través de la cría de animales , lo que los haría "inferiores" en términos de selección natural .
Bases genéticas y epigenéticas
Desde principios de la década de 1900, se han desarrollado dos hipótesis genéticas en competencia, no necesariamente mutuamente excluyentes, para explicar el vigor híbrido. Más recientemente, también se ha establecido un componente epigenético de vigor híbrido. [11] [12]
La hipótesis de dominancia genética atribuye la superioridad de los híbridos al enmascaramiento de la expresión de alelos recesivos indeseables (deletéreos) de un padre por alelos dominantes (generalmente de tipo salvaje) del otro (ver Complementación (genética) ). Atribuye el bajo rendimiento de las cepas consanguíneas a la expresión de alelos recesivos deletéreos homocigotos. La hipótesis de sobredominio genético establece que algunas combinaciones de alelos (que pueden obtenerse cruzando dos cepas puras) son especialmente ventajosas cuando se emparejan en un individuo heterocigoto . Esta hipótesis se invoca comúnmente para explicar la persistencia de algunos alelos (el más famoso es el rasgo de células falciformesalelo) que son dañinos en homocigotos. En circunstancias normales, estos alelos dañinos se eliminarían de una población mediante el proceso de selección natural. Al igual que la hipótesis de dominancia, atribuye el bajo rendimiento de las cepas consanguíneas a la expresión de alelos recesivos tan dañinos. En cualquier caso, los cruces cruzados proporcionan el beneficio de enmascarar los alelos recesivos deletéreos en la progenie. Se ha propuesto que este beneficio es un factor importante en el mantenimiento de la reproducción sexual entre eucariotas, como se resume en el artículo Evolución de la reproducción sexual .
Se ha establecido una contribución epigenética a la heterosis en plantas, [12] y también se ha informado en animales. [13] Los microARN (miARN), descubiertos en 1993, son una clase de ARN pequeños no codificantes que reprimen la traducción de los ARN mensajeros (ARNm) o provocan la degradación de los ARNm. [14] En las plantas híbridas, la mayoría de los miARN tienen una expresión no aditiva (puede ser mayor o menor que los niveles en los padres). [12] Esto sugiere que los ARN pequeños están involucrados en el crecimiento, vigor y adaptación de los híbridos. [12]
Los efectos de la 'heterosis sin hibridación' sobre el tamaño de la planta se han demostrado en plantas triploides (autopoliploides) F1 genéticamente isogénicas, donde los triploides F1 en exceso del genoma paterno muestran heterosis positiva, mientras que los F1 en exceso del genoma materno muestran efectos de heterosis negativos. [15] Estos hallazgos demuestran que los efectos de heterosis, con una base epigenética dependiente de la dosis del genoma, pueden generarse en la descendencia F1 que son genéticamente isogénicos (es decir, no albergan heterocigosidad). [15] [16] Se ha demostrado [11] que el vigor híbrido en un híbrido alopoliploide de dos Arabidopsisespecie se debió al control epigenético en las regiones aguas arriba de dos genes, lo que provocó una alteración importante aguas abajo en la acumulación de clorofila y almidón. El mecanismo implica la acetilación y / o metilación de aminoácidos específicos en la histona H3 , una proteína estrechamente asociada con el ADN, que puede activar o reprimir genes asociados.
Complejo mayor de histocompatibilidad en animales
Un ejemplo de dónde genes particulares pueden ser importantes en animales vertebrados para la heterosis es el complejo principal de histocompatibilidad (MHC). Los vertebrados heredan varias copias de MHC de clase I y MHC de clase II de cada padre, que se utilizan en la presentación de antígenos como parte del sistema inmunológico adaptativo. Cada copia diferente de los genes puede unirse y presentar un conjunto diferente de péptidos potenciales a los linfocitos T. Estos genes son altamente polimórficos en todas las poblaciones, pero son más similares en poblaciones más pequeñas y más estrechamente relacionadas. La reproducción entre individuos más distantes genéticamente disminuye la posibilidad de heredar dos alelos iguales o similares, lo que permite presentar una gama más diversa de péptidos. Esto, por lo tanto, reduce la posibilidad de que no se reconozca un patógeno en particular, y significa que es probable que se reconozcan más proteínas antigénicas en cualquier patógeno, lo que da un mayor rango de activación de las células T y, por lo tanto, una mayor respuesta. Esto también significa que la inmunidad adquirida al patógeno es contra una mayor variedad de antígenos, lo que significa que el patógeno debe mutar.más antes de que se pierda la inmunidad. Por lo tanto, los híbridos tienen menos probabilidades de sucumbir a enfermedades patógenas y son más capaces de combatir las infecciones. Sin embargo, esta puede ser la causa de enfermedades autoinmunes. [ cita requerida ]
Plantas
Los cruces entre purasangre de diferentes grupos heteróticos dan como resultado híbridos F1 vigorosos con una heterosis significativamente mayor que los híbridos F1 de purasangre dentro del mismo grupo o patrón heterótico. Los fitomejoradores crean grupos heteróticos para clasificar las líneas puras y pueden mejorarse progresivamente mediante la selección recíproca recurrente.
La heterosis se utiliza para aumentar los rendimientos, la uniformidad y el vigor. Los métodos de cría híbridos se utilizan en maíz , sorgo , arroz , remolacha azucarera , cebolla , espinaca , girasoles , brócoli y para crear un cannabis más psicoactivo .
Maíz (maíz)
Casi todo el maíz de campo ( maíz ) cultivado en la mayoría de las naciones desarrolladas exhibe heterosis. Los híbridos de maíz modernos superan sustancialmente el rendimiento de los cultivares convencionales y responden mejor a los fertilizantes .
La heterosis del maíz fue célebremente demostrada a principios del siglo XX por George H. Shull y Edward M. East después de que el Dr. William James Beal, de la Universidad Estatal de Michigan, inventara el maíz híbrido, basándose en un trabajo iniciado en 1879 a instancias de Charles Darwin . El trabajo del Dr. Beal condujo al primer relato publicado de un experimento de campo que demuestra el vigor híbrido en el maíz, por Eugene Davenport y Perry Holden., 1881. Estos diversos pioneros de la botánica y campos relacionados demostraron que los cruces de líneas puras hechas de una abolladura del sur y un pedernal del norte, respectivamente, mostraron una heterosis sustancial y superaron el rendimiento de los cultivares convencionales de esa época. Sin embargo, en ese momento esos híbridos no podían fabricarse económicamente a gran escala para que los usaran los agricultores. Donald F. Jones en la Estación Experimental Agrícola de Connecticut , New Haveninventó el primer método práctico para producir un maíz híbrido de alto rendimiento en 1914-1917. El método de Jones produjo un híbrido de doble cruzamiento, que requiere dos pasos de cruzamiento que trabajan a partir de cuatro líneas endogámicas originales distintas. El trabajo posterior de los mejoradores de maíz produjo líneas endogámicas con suficiente vigor para la producción práctica de un híbrido comercial en un solo paso, los híbridos de un solo cruce. Los híbridos de un solo cruzamiento se hacen a partir de solo dos padres endogámicos originales. Por lo general, son más vigorosos y también más uniformes que los híbridos de doble cruzamiento anteriores. El proceso de creación de estos híbridos a menudo implica el desespillado .
Los híbridos de maíz templado se derivan de dos grupos heteróticos principales: 'Iowa Stiff Stalk Synthetic' y tallo no rígido. [ cita requerida ]
Arroz ( Oryza sativa )
La producción de arroz ha experimentado un enorme aumento en China debido al uso intensivo de arroz híbrido. En China, los esfuerzos han generado una variedad de arroz súper híbrido ('LYP9') con una capacidad de producción de alrededor de 15 toneladas por hectárea. También en la India, varias variedades han mostrado un gran vigor, incluidas 'RH-10' y 'Suruchi 5401'.
Ganado híbrido
El concepto de heterosis también se aplica en la producción de ganado comercial . En el ganado, los cruces entre Black Angus y Hereford producen un cruce conocido como " Black Baldy ". En los cerdos , las "colillas azules" son producidas por el cruce de Hampshire y Yorkshire. Otros híbridos más exóticos (dos especies diferentes, por lo que genéticamente son más diferentes), como el " beefalo ", también se utilizan para los mercados de especialidades.
Aves de corral
Dentro de las aves de corral , se han utilizado genes ligados al sexo para crear híbridos en los que los machos y las hembras pueden clasificarse por color al día de edad. Los genes específicos utilizados para esto son genes para el crecimiento de las plumas de las alas y las barreras. Las cruces de este tipo crean lo que se venden como enlaces sexuales negros, enlaces sexuales rojos y varios otros cruces que se conocen por nombres comerciales.
Los pollos de engorde comerciales se producen cruzando diferentes cepas de White Rocks y White Cornish, el Cornish proporciona un marco grande y el Rocks proporciona una rápida tasa de ganancia. El vigor híbrido producido permite la producción de aves uniformes con un peso de canal comercializable a las 6-9 semanas de edad.
Asimismo, los híbridos entre diferentes cepas de White Leghorn se utilizan para producir bandadas ponedoras que proporcionan la mayoría de los huevos blancos para la venta en los Estados Unidos.
Perros
En 2013, un estudio encontró que las razas mixtas viven en promedio 1,2 años más que las razas puras. [17]
John Scott y John L. Fuller realizaron un estudio detallado de Cocker Spaniels de pura raza, Basenjis de pura raza e híbridos entre ellos. [18] Descubrieron que los híbridos corrían más rápido que cualquiera de los padres, quizás debido a la heterosis. Otras características, como la frecuencia cardíaca basal, no mostraron heterosis (la frecuencia cardíaca basal del perro estaba cerca del promedio de sus padres), quizás debido a los efectos aditivos de múltiples genes. [19]
A veces, las personas que trabajan en un programa de cría de perros no encuentran una heterosis útil. [20]
Dicho todo esto, los estudios no proporcionan una prueba definitiva del vigor híbrido en perros. Esto se debe en gran parte a la herencia desconocida de la mayoría de los perros mestizos que se utilizan. Los resultados varían enormemente, con algunos estudios que muestran beneficios y otros encuentran que los perros de razas mixtas son más propensos a las condiciones genéticas. [21] [22] [23]
Aves
En 2014, un estudio realizado por el Centro de Ecología Integrativa de la Universidad Deakin en Geelong, Victoria, concluyó que los híbridos intraespecíficos entre la subespecie Platycercus elegans flaveolus y P. e. elegans de la rosella carmesí ( P. elegans ) tenían más probabilidades de combatir enfermedades que sus contrapartes puras. [24]
Humanos
Los seres humanos son todos extremadamente similares genéticamente entre sí. [25] [26] [27] Michael Mingroni ha propuesto la heterosis, en forma de vigor híbrido asociado con reducciones históricas de los niveles de endogamia, como una explicación del efecto Flynn , el aumento constante en los puntajes de las pruebas de CI en todo el mundo durante el siglo 20.
Ver también
- F1 híbrido
- Mezcla genética
- Ventaja heterocigota
Referencias
- ^ George Harrison Shull (1948). "¿Qué es la" heterosis "?" . Genética . 33 (5): 439–446. PMC 1209417 . PMID 17247290 .
- ^ Birchler JA; Auger DL; Acertijo NC (2003). "En busca de la base molecular de la heterosis" . La célula vegetal . 15 (10): 2236–2239. doi : 10.1105 / tpc.151030 . PMC 540269 . PMID 14523245 .
- ^ Cuervo, James F. (1948). "Hipótesis alternativas de vigor híbrido" . Genética . 33 (5): 477–487. PMC 1209419 . PMID 17247292 .
- ^ Davenport CB (1908). "Degeneración, albinismo y endogamia" . Ciencia . 28 (718): 454–5. Código Bibliográfico : 1908Sci .... 28..454D . doi : 10.1126 / science.28.718.454-b . PMID 17771943 .
- ^ a b Carr, David E .; Dudash, Michele R. (29 de junio de 2003). "Enfoques recientes sobre la base genética de la depresión endogámica en las plantas" . Transacciones filosóficas de la Royal Society B: Ciencias biológicas . 358 (1434): 1071–1084. doi : 10.1098 / rstb.2003.1295 . ISSN 0962-8436 . PMC 1693197 . PMID 12831473 .
- ↑ East EM (1908). "Consanguinidad en maíz". Informes de la estación de experimentos agrícolas de Connecticut para 1907 : 419–428.
- ^ Shull GH (1908). "La composición de un campo de maíz". Informes de la Asociación Estadounidense de Criadores : 296–301.
- ↑ a b Crow, James F. (1998). "Hace 90 años: el comienzo del maíz híbrido" . Genética . 148 (3): 923–928. PMC 1460037 . PMID 9539413 .
- ↑ a b Risch N, Burchard E, Ziv E, Tang H (julio de 2002). "Categorización de los seres humanos en la investigación biomédica: genes, raza y enfermedad" . Genome Biol . 3 (7): comentario 2007. doi : 10.1186 / gb-2002-3-7-comment2007 . PMC 139378 . PMID 12184798 .
- ^ Weller SG, Sakai AK, Thai DA, Tom J, Rankin AE (noviembre de 2005). "Depresión endogámica y heterosis en poblaciones de Schiedea viscosa, una especie altamente autofecundada" . J. Evol. Biol . 18 (6): 1434–44. doi : 10.1111 / j.1420-9101.2005.00965.x . PMID 16313456 .
- ^ a b Ni Z, Kim ED, Ha M, et al. (Enero de 2009). "Los ritmos circadianos alterados regulan el vigor del crecimiento en híbridos y alopoliploides" . Naturaleza . 457 (7227): 327–31. Código Bibliográfico : 2009Natur.457..327N . doi : 10.1038 / nature07523 . PMC 2679702 . PMID 19029881 .
- ↑ a b c d Baranwal VK, Mikkilineni V, Zehr UB, Tyagi AK, Kapoor S (noviembre de 2012). "Heterosis: ideas emergentes sobre el vigor híbrido" . J. Exp. Bot . 63 (18): 6309-14. doi : 10.1093 / jxb / ers291 . PMID 23095992 .
- ^ Han Z, Mtango NR, Patel BG, Sapienza C, Latham KE (octubre de 2008). "Vigor híbrido y efectos epigenéticos transgeneracionales en el fenotipo de embrión de ratón temprano" . Biol. Reprod . 79 (4): 638–48. doi : 10.1095 / biolreprod.108.069096 . PMC 2844494 . PMID 18562704 .
- ^ Zhou Y, Ferguson J, Chang JT, Kluger Y (2007). "Regulación inter e intra-combinatoria por factores de transcripción y microARN" . BMC Genomics . 8 : 396. doi : 10.1186 / 1471-2164-8-396 . PMC 2206040 . PMID 17971223 .
- ^ a b Fuerte, Antoine; Ryder, Peter; McKeown, Peter C .; Wijnen, Cris; Aarts, Mark G .; Sulpice, Ronan; Spillane, Charles (1 de enero de 2016). "Desagregación de poliploidía, dosificación del genoma parental y contribuciones de hibridación a la heterosis en Arabidopsis thaliana" . El nuevo fitólogo . 209 (2): 590–599. doi : 10.1111 / nph.13650 . ISSN 1469-8137 . PMID 26395035 .
- ^ Duszynska, Dorota; McKeown, Peter C .; Juenger, Thomas E .; Pietraszewska-Bogiel, Anna; Geelen, Danny; Spillane, Charles (1 de abril de 2013). "La fertilidad de gametos y la variación del número de óvulos en plantas triploides híbridas F1 recíprocas autofecundadas son hereditarias y muestran efectos epigenéticos de origen" . El nuevo fitólogo . 198 (1): 71–81. doi : 10.1111 / nph.12147 . ISSN 1469-8137 . PMID 23368793 .
- ^ O'Neill, DG; Church, DB; McGreevy, PD; Thomson, PC; Brodbelt, DC (2013). "Longevidad y mortalidad de los perros de propiedad en Inglaterra" (PDF) . La Revista Veterinaria . 198 (3): 638–43. doi : 10.1016 / j.tvjl.2013.09.020 . PMID 24206631 .
- ^ Spady, Tyrone C .; Ostrander, Elaine A. (2008). "Genética del comportamiento canino: señalar los fenotipos y reunir los genes" . Revista Estadounidense de Genética Humana . 82 : 10–8. doi : 10.1016 / j.ajhg.2007.12.001 . PMC 2253978 . PMID 18179880 .
- ^ John Paul Scott y John L. Fuller. "Genética y comportamiento social del perro" . 1965. pág. 307 y p. 313.
- ^ Per Jensen. "La biología del comportamiento de los perros" . 2007. p. 179
- ^ Nicolás, Frank W (2016). "¿Vigor híbrido en perros?" . Pubmed . Biblioteca Nacional de Medicina . Consultado el 29 de julio de 2020 .
- ^ Nicolás, JAC (2012). "Encuesta de anomalías oftálmicas en el labradoodle en el Reino Unido" . Pubmed . Biblioteca Nacional de Medicina . Consultado el 29 de julio de 2020 .
- ^ Sharkey, Laura (2020). "Vigor híbrido en perros" . The Functional Dog Collaborative . The Functional Dog Collaborative . Consultado el 29 de julio de 2020 .
- ^ Australian Geographic (septiembre de 2014). "Las aves híbridas son mejores para combatir las enfermedades que las razas puras" .
- ^ Halcones, John (2013). Importancia de los genomas neandertales y denisovanos en la evolución humana . Revisión anual de antropología . 42 . Revisiones anuales. págs. 433–449, 438. doi : 10.1146 / annurev-anthro-092412-155548 . ISBN 978-0-8243-1942-7. ISSN 0084-6570 .
La historia evolutiva compartida de los seres humanos vivos ha resultado en una alta relación entre todas las personas vivas, como lo indica, por ejemplo, el índice de fijación muy bajo (F ST ) entre las poblaciones humanas vivas.
- ^ Barbujani, Guido ; Colonna, Vincenza (15 de septiembre de 2011). "Capítulo 6: Base genética de la biodiversidad humana: una actualización" . En Zachos, Frank E .; Habel, Jan Christian (eds.). Puntos críticos de biodiversidad: distribución y protección de áreas prioritarias de conservación . Saltador. págs. 97-119. doi : 10.1007 / 978-3-642-20992-5_6 . ISBN 978-3-642-20992-5. Consultado el 23 de noviembre de 2013 .
Los esfuerzos masivos para estudiar el genoma humano en detalle han producido cantidades extraordinarias de datos genéticos. Aunque todavía no comprendemos las bases moleculares de la mayoría de los rasgos complejos, incluidas muchas enfermedades comunes, ahora tenemos una idea más clara del grado de semejanza genética entre los humanos y otras especies de primates. También sabemos que los seres humanos están genéticamente muy cerca unos de otros, de hecho más que cualquier otro primate, que la mayor parte de nuestra diversidad genética se explica por diferencias individuales dentro de las poblaciones, y que solo una pequeña fracción de la variación genética de las especies se encuentra entre poblaciones. y grupos geográficos de los mismos.
- ^ Ramachandran, Sohini; Tang, Hua; Gutenkunst, Ryan N .; Bustamante, Carlos D. (2010). "Capítulo 20: Genética y genómica de la estructura de la población humana" (PDF) . En Speicher, Michael R .; Antonarakis, Stylianos E .; Motulsky, Arno G. (eds.). Genética humana de Vogel y Motulsky: problemas y enfoques . Heidelberg: Springer Scientific. págs. 589–615. doi : 10.1007 / 978-3-540-37654-5 . ISBN 978-3-540-37653-8. Archivado desde el original (PDF) el 3 de diciembre de 2013 . Consultado el 29 de octubre de 2013 . Lay resumen (4 de septiembre de 2010).
La mayoría de los estudios de genética de poblaciones humanas comienzan citando un artículo seminal de 1972 de Richard Lewontin que lleva el título de esta subsección [29]. Dado el papel central que este trabajo ha jugado en nuestro campo, comenzaremos por discutirlo brevemente y regresaremos a sus conclusiones a lo largo del capítulo. ... Una conclusión clave del artículo es que el 85,4% de la variación genética total observada ocurrió dentro de cada grupo. Es decir, informó que la gran mayoría de las diferencias genéticas se encuentran dentro de las poblaciones y no entre ellas. ... Su hallazgo se ha reproducido en estudio tras estudio hasta el presente: dos individuos aleatorios de cualquier grupo (que podría ser un continente o incluso una población local) son casi tan diferentes como dos individuos aleatorios de todo el mundo.
Lectura adicional
- Bakker, Winfridus (2006). "El vigor híbrido mejorado beneficia a reproductoras y pollos de engorde" (PDF) . Enfoque Cobb (2). Archivado desde el original (PDF) el 17 de diciembre de 2008.
- Birchler JA, Auger DL, Riddle NC (octubre de 2003). "En busca de la base molecular de la heterosis" . Célula vegetal . 15 (10): 2236–9. doi : 10.1105 / tpc.151030 . PMC 540269 . PMID 14523245 .
- NOAA Tech Memo NMFS NWFSC-30: Efectos genéticos de la desviación de peces de criadero no nativos en poblaciones naturales: depresión por consanguinidad y depresión por consanguinidad
- Mac Gregor, SE (2009) [1976]. "Introducción: vigor híbrido en plantas y su relación con la polinización de insectos" (PDF) . Polinización por insectos de plantas cultivadas . Manual de agricultura no. 496. Servicio de Investigación Agrícola, USDA OCLC 243509268 .
- "Híbridos y reliquias": un artículo de Home Hort Hints de University of Illinois Extension
- Roybal, J. (1 de julio de 1998). " Ranchstar ". Carne de res (beefmagazine.com).
- "Sex-Links": en relación con las aves de corral; en FeatherSite