| Pterosaurios | |
|---|---|
 | |
| Réplica de esqueletos de Geosternbergia sternbergi , femenino (izquierda) y masculino (derecha) | |
| clasificación cientifica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Clade : | Ornithodira |
| Clade : | † Pterosauromorpha |
| Orden: | † Pterosauria Kaup , 1834 |
| Subgrupos [1] [2] | |
| |
 | |
| Distribución de ubicaciones de fósiles de pterosaurios . Los nombres de especies o géneros coloreados corresponden a su grupo taxonómico. [a] | |
Pterosaurs ( / t ɛr ə s ɔr , t ɛr oʊ - / ; [5] [6] del griego pteron y sauros , que significa "lagarto ala") estaban volando reptiles de la extinta clado o fin Pterosauria . Existieron durante la mayor parte del Mesozoico : desde finales del Triásico hasta finales del Cretácico ( hace 228 a 66 millones de años [7] ). Los pterosaurios son los primeros vertebradosconocido por haber evolucionado el vuelo motorizado . Sus alas estaban formadas por una membrana de piel, músculos y otros tejidos que se extendían desde los tobillos hasta un dedo anular dramáticamente alargado. [8]
Había dos tipos principales de pterosaurios. Los pterosaurios basales (también llamados 'pterosaurios no pterodactiloides' o 'rhamphorhynchoids') eran animales más pequeños con mandíbulas completamente dentadas y, típicamente, colas largas. Las anchas membranas de sus alas probablemente incluían y conectaban las patas traseras. En el suelo, habrían tenido una postura incómoda y tendida, pero la anatomía de sus articulaciones y sus fuertes garras los habrían convertido en trepadores efectivos, y es posible que hayan vivido en los árboles. Los pterosaurios basales eran insectívoros o depredadores.de pequeños vertebrados. Los pterosaurios posteriores (pterodactiloides) desarrollaron muchos tamaños, formas y estilos de vida. Los pterodactiloides tenían alas más estrechas con extremidades posteriores libres, colas muy reducidas y cuellos largos con cabezas grandes. En el suelo, los pterodactiloides caminaban bien sobre las cuatro extremidades con una postura erguida, de pie plantígrados sobre las patas traseras y doblando el dedo del ala hacia arriba para caminar sobre la "mano" de tres dedos. Podían despegar del suelo, y los rastros fósiles muestran que al menos algunas especies pudieron correr, vadear o nadar. [9] Sus mandíbulas tenían picos córneos y algunos grupos carecían de dientes. Algunos grupos desarrollaron crestas elaboradas en la cabeza con dimorfismo sexual .
Los pterosaurios lucían capas de filamentos similares a pelos conocidos como picnofibras , que cubrían sus cuerpos y partes de sus alas. Las picnofibras crecieron en varias formas, desde simples filamentos hasta plumas ramificadas. Estas son posiblemente homólogas a las plumas que se encuentran tanto en los dinosaurios aviares como en algunos no aviares, lo que sugiere que las primeras plumas evolucionaron en el antepasado común de los pterosaurios y los dinosaurios, posiblemente como aislante. [10] En vida, los pterosaurios habrían tenido un pelaje suave o esponjoso que no se parecía a las plumas de las aves. Eran animales activos de sangre caliente (endotérmicos). El sistema respiratorio tenía una respiración de "flujo continuo" unidireccional eficiente utilizando sacos de aire., que ahuecó sus huesos hasta un extremo. Los pterosaurios abarcaron una amplia gama de tamaños adultos, desde los anurognátidos muy pequeños hasta las criaturas voladoras más grandes conocidas de todos los tiempos, incluidos Quetzalcoatlus y Hatzegopteryx , [11] [12] [13] que alcanzaron una envergadura de al menos nueve metros. La combinación de endotermia , un buen suministro de oxígeno y músculos fuertes hicieron a los pterosaurios voladores poderosos y capaces.
Los pterosaurios son a menudo referidos por los medios populares o el público en general como " dinosaurios voladores ", pero los dinosaurios se definen como los descendientes del último ancestro común de Saurischia y Ornithischia , que excluye a los pterosaurios. [14] No obstante, los pterosaurios están más estrechamente relacionados con las aves y otros dinosaurios que con los cocodrilos o cualquier otro reptil viviente, aunque no son antepasados de las aves. Los pterosaurios también se conocen coloquialmente como pterodáctilos , particularmente en la ficción y el periodismo. [15] Sin embargo, técnicamente, pterodactyl solo se refiere a miembros del género Pterodactylus, y más ampliamente a los miembros del suborden Pterodactyloidea de los pterosaurios. [dieciséis]
Los pterosaurios tenían una variedad de estilos de vida. Tradicionalmente visto como comedores de pescado, ahora se entiende que el grupo incluía cazadores de animales terrestres, insectívoros, comedores de frutas e incluso depredadores de otros pterosaurios. Se reprodujeron mediante huevos, de los cuales se han descubierto algunos fósiles.
Descripción
La anatomía de los pterosaurios fue muy modificada de sus antepasados reptiles por la adaptación al vuelo. Los huesos de pterosaurio eran huecos y estaban llenos de aire, como los de los pájaros . Esto proporcionó una superficie de inserción muscular más alta para un peso esquelético dado. Las paredes óseas eran a menudo delgadas como el papel. Tenían un esternón grande y con quilla para los músculos de vuelo y un cerebro agrandado capaz de coordinar un comportamiento de vuelo complejo. [17] Los esqueletos de pterosaurios a menudo muestran una fusión considerable. En el cráneo desaparecieron las suturas entre elementos. En algunos pterosaurios posteriores, la columna vertebral sobre los hombros se fusionó en una estructura conocida como notario, que sirvió para endurecer el torso durante el vuelo y proporcionar un soporte estable para el omóplato . Del mismo modo, las vértebras sacras podrían formar un solo synsacro mientras que los huesos pélvicos también se fusionaron.
Los pterosaurios basales incluyen los clados Dimorphodontidae ( Dimorphodon ), Campylognathididae ( Eudimorphodon , Campyognathoides ) y Rhamphorhynchidae ( Rhamphorhynchus , Scaphognathus ).
Pterodactyloids incluyen los clados ornithocheiroidea ( istiodactylus , Ornithocheirus , Pteranodon ), ctenochasmatoidea ( ctenochasma , Pterodactylus ), Dsungaripteroidea ( germanodactylus , Dsungaripterus ), y azhdarchoidea ( Tapejara , Tupuxuara , Quetzalcoatlus ).
Los dos grupos se superpusieron en el tiempo, pero los primeros pterosaurios en el registro fósil son los pterosaurios basales, y los últimos pterosaurios son los pterodactiloides. [18]
Se debate la posición del clado Anurognathidae ( Anurognathus , Jeholopterus , Vesperopterylus ). [19] Los anurognátidos (pterosaurios con cabeza de rana) eran altamente especializados. Voladores pequeños con mandíbulas acortadas y una gran abertura, algunos tenían ojos grandes que sugerían hábitos nocturnos o crepusculares , cerdas en la boca y pies adaptados para agarrarse. Se observan adaptaciones paralelas en aves y murciélagos que se alimentan de insectos en vuelo.
Tamaño
Los pterosaurios tenían una amplia gama de tamaños. Generalmente eran bastante grandes. Incluso las especies más pequeñas tenían una envergadura de no menos de 25 centímetros (10 pulgadas). [11] Las formas más grandes representan los animales más grandes que jamás hayan volado, con envergaduras de hasta 10-11 metros (33-36 pies). [20]
De pie, estos gigantes podrían alcanzar la altura de una jirafa moderna . Tradicionalmente, se suponía que los pterosaurios eran extremadamente ligeros en relación con su tamaño. Más tarde, se entendió que esto implicaría densidades poco realistas de sus tejidos blandos. Por lo tanto, algunas estimaciones modernas extrapolan un peso de hasta 250 kilogramos (550 libras) para las especies más grandes. [21]
Cráneo, dientes y crestas
En comparación con los otros grupos voladores de vertebrados, las aves y los murciélagos, los cráneos de pterosaurios eran típicamente bastante grandes. [22] La mayoría de los cráneos de pterosaurios tenían mandíbulas alargadas. [22] Los huesos del cráneo tienden a fusionarse en individuos adultos. [22] Los primeros pterosaurios a menudo tenían dientes heterodontes , de constitución variable, y algunos todavía tenían dientes en el paladar. En grupos posteriores, la mayoría de los dientes se volvieron cónicos. [23] Los dientes frontales a menudo eran más largos, formando un "agarre de presa" en las puntas de las mandíbulas expandidas transversalmente, pero el tamaño y la posición eran muy variables entre las especies. [24] Con la Pterodactyloidea derivada, los cráneos se alargaron aún más, a veces sobrepasando en longitud el cuello y el torso combinados. Esto fue causado por un estiramiento y fusión del hueso frontal del hocico, el premaxilar , con el hueso de la mandíbula superior, el maxilar . A diferencia de la mayoría de los arcosaurios , las aberturas nasal y antorbitaria de los pterosaurios pterodactiloides se fusionaron en una única abertura grande, llamada fenestra nasoantorbital . [25] Esta característica probablemente evolucionó para aligerar el cráneo para volar. [23] En contraste, los huesos detrás de la cuenca del ojo se contrajeron y rotaron, inclinando fuertemente la parte posterior del cráneo y llevando la articulación de la mandíbula hacia adelante. [26] La caja del cerebroera relativamente grande para los reptiles. [27]
En algunos casos, se ha conservado el tejido del pico queratinoso fosilizado , aunque en formas dentadas, el pico es pequeño y se limita a las puntas de la mandíbula y no afecta a los dientes. [28] Algunas formas avanzadas con pico no tenían dientes, como los Pteranodontidae y Azhdarchidae , y tenían picos más grandes, más extensos y más parecidos a los de las aves. [23] Algunos grupos tenían formas dentales especializadas. Los Istiodactylidae tenían dientes curvados para comer carne. Ctenochasmatidae utilizó peines de numerosos dientes en forma de aguja para la alimentación por filtración; Pterodaustro podría tener más de mil dientes con forma de cerdas. Dsungaripteridaecubrieron sus dientes con tejido de la mandíbula para una función de aplastamiento. Si había dientes, se colocaron en cavidades dentales separadas. [25] Los dientes de reemplazo se generaron detrás, no debajo, de los dientes más viejos. [24]
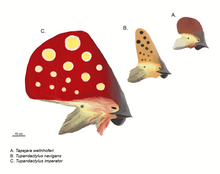
La imagen pública de los pterosaurios se define por sus elaboradas crestas en la cabeza. [29] Esto fue influenciado por la distintiva cresta apuntando hacia atrás del conocido Pteranodon . Las posiciones principales de tales crestas son la parte frontal del hocico, como una excrecencia de los premaxilares, o la parte posterior del cráneo como una extensión de los huesos parietales, en cuyo caso se denomina "cresta supraoccipital". [27] Las crestas delanteras y traseras pueden estar presentes simultáneamente y pueden fusionarse en una sola estructura más grande, la más expansiva de las cuales es la que muestran los Tapejaridae . Nyctosauruslucía una extraña cresta en forma de cuerno. Las crestas tenían solo unos pocos milímetros de grosor transversalmente. La base de la cresta ósea típicamente estaría extendida por tejido queratinoso u otro tejido blando. [27]
Desde la década de 1990, nuevos descubrimientos y un estudio más completo de especímenes antiguos han demostrado que las crestas están mucho más extendidas entre los pterosaurios de lo que se suponía anteriormente. El hecho de que estuvieran extendidos o compuestos completamente de queratina, que no se fosiliza fácilmente, había inducido a error a investigaciones anteriores. [30] Para Pterorhynchus y Pterodactylus , la verdadera extensión de estas crestas solo se ha descubierto mediante fotografía ultravioleta . [28] [31] Mientras que las crestas fósiles solían estar restringidas a los Pterodactyloidea más avanzados, Pterorhynchus y Austriadactylus muestran que incluso algunos pterosaurios primitivos las poseían. [30]
Al igual que las mandíbulas superiores, las mandíbulas inferiores emparejadas de los pterosaurios eran muy alargadas. [32] En las formas avanzadas, tendían a ser más cortos que la parte superior del cráneo porque la articulación de la mandíbula estaba en una posición más adelantada. Los huesos delanteros de la mandíbula inferior, los dentarios u ossa dentalia , estaban en la punta fusionados firmemente en una sínfisis central. Esto hizo que las mandíbulas inferiores funcionaran como un todo conectado, la mandíbula . La sínfisis era a menudo muy delgada transversalmente y larga, representando una parte considerable de la longitud de la mandíbula, hasta el 60%. [26] Si hubiera una cresta en el hocico, la sínfisis podría presentar una cresta mandibular a juego, sobresaliendo hacia abajo. [26]Las especies dentadas también tienen dientes en sus dentarios. La mandíbula se abría y cerraba con un simple movimiento vertical u "ortogonal" hacia arriba y hacia abajo.
Columna vertebral
La columna vertebral de los pterosaurios contaba entre treinta y cuatro y setenta vértebras . Las vértebras delante de la cola eran "procelulares": el cotilo (frente del cuerpo vertebral ) era cóncavo y en él encajaba una extensión convexa en la parte posterior de la vértebra anterior, el cóndilo . Los pterosaurios avanzados son únicos en poseer procesos especiales que se proyectan adyacentes a su cóndilo y cotilo, los exapófisis , [33] y el cotilo también puede poseer una pequeña punta en su línea media llamada hipopófisis. [34]
El cuello de los pterosaurios era relativamente largo y recto. En los pterodactiloides, el cuello suele ser más largo que el torso. [35] Esta longitud no se debe a un aumento del número de vértebras, que invariablemente es siete. Algunos investigadores incluyen dos "cervicodorsales" de transición, lo que eleva el número a nueve. [35] En cambio, las vértebras mismas se alargaron más, hasta ocho veces más largas que anchas. Sin embargo, las cervicales eran más anchas que altas, lo que implicaba una mejor movilidad vertical del cuello que horizontal. Los pterodactiloides han perdido todas las costillas del cuello. [34] Los cuellos de los pterosaurios probablemente eran bastante gruesos y bien musculosos, [36] especialmente en vertical. [37]
El torso era relativamente corto y en forma de huevo. Las vértebras en la parte posterior de los pterosaurios originalmente podrían haber numerado dieciocho. Con las especies avanzadas, un número creciente de estas tendía a incorporarse al sacro . Estas especies también muestran a menudo una fusión de las vértebras dorsales anteriores en un todo rígido que se llama notario por una estructura comparable en las aves. Esta fue una adaptación para resistir las fuerzas causadas por batir las alas. [35] El notario incluía de tres a siete vértebras, dependiendo de la especie involucrada, pero también de la edad del individuo. Estas vértebras podrían estar conectadas por tendones o una fusión de sus espinas neurales en una "placa supraneural". Sus costillas también se fusionarían firmemente en el notario.[38] En general, las costillas son de dos cabezas. [39] El sacro constaba de tres a diez vértebras sacras. Ellos también podrían conectarse a través de una placa supraneural que, sin embargo, no entraría en contacto con el notario. [38]
Las colas de los pterosaurios siempre fueron bastante delgadas. Esto significa que el músculo retractor caudofemoralis , que en la mayoría de Archosauria basal proporciona la principal fuerza propulsora de la extremidad trasera, era relativamente poco importante. [37] Las vértebras de la cola eran anfícolas y los cuerpos vertebrales de ambos extremos eran cóncavos. Las primeras especies tenían colas largas, que contenían hasta cincuenta vértebras caudales, las del medio estaban rígidas por procesos de articulación alargados, las cigapófisis y los galones . [40] Tales colas actuaban como timones, a veces terminando en la parte trasera en una paleta vertical en forma de diamante u ovalada. [41] En los pterodactiloides, las colas se redujeron mucho y nunca se endurecieron,[41] con algunas especies contando hasta diez vértebras. [38]
Cintura escapular
La cintura escapular era una estructura fuerte que transfirió las fuerzas del vuelo aleteando al tórax . Probablemente estaba cubierto por gruesas capas musculares. [42] El hueso superior, el omóplato , era una barra recta. Estaba conectado a un hueso inferior, el coracoide que es relativamente largo en los pterosaurios. En las especies avanzadas, su conjunto combinado, el escapulocoracoideo, estaba orientado casi verticalmente. En ese caso, el omóplato encajaba en un hueco en el costado del notario, mientras que el coracoides también se conectaba al esternón. De esta manera, ambos lados juntos formaron un circuito cerrado rígido, capaz de soportar fuerzas considerables. [39]Una peculiaridad era que las conexiones del esternón de los coracoides a menudo eran asimétricas, con un coracoides unido delante del otro. En especies avanzadas, la articulación del hombro se había movido desde el omóplato hasta la coracoides. [43] La articulación tenía forma de silla de montar y permitía un movimiento considerable del ala. [39] Estaba orientado hacia los lados y algo hacia arriba. [41]
El esternón, formado por esternones emparejados fusionados , era ancho. Tenía solo una quilla poco profunda. A través de las costillas del esternón, estaba unido a sus lados a las costillas dorsales. [40] En su parte posterior, una fila de costillas del vientre o gastralia estaba presente, cubriendo todo el vientre. [41] Al frente, una punta larga, la cristospina , sobresalía oblicuamente hacia arriba. El borde posterior del esternón era el punto más profundo del tórax. [43] Las clavículas o las interclavículas estaban completamente ausentes. [41]
Alas
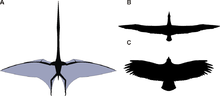
Las alas de los pterosaurios estaban formadas por huesos y membranas de piel y otros tejidos. Las membranas primarias se adhieren al dedo anular extremadamente largo de cada brazo y se extienden a lo largo de los lados del cuerpo. Dónde terminaron ha sido muy controvertido, pero desde la década de 1990 se han encontrado una docena de especímenes con tejidos blandos conservados que parecen mostrar adheridos a los tobillos. [44] Sin embargo, la curvatura exacta del borde de fuga sigue siendo equívoca. [44]
Si bien históricamente se ha considerado como simples estructuras correosas compuestas de piel, la investigación ha demostrado desde entonces que las membranas de las alas de los pterosaurios eran estructuras dinámicas muy complejas adecuadas para un estilo de vuelo activo. [45] Las alas externas (desde la punta hasta el codo) fueron reforzadas por fibras muy espaciadas llamadas actinofibrillas . [46] Las actinofibrillas mismas constaban de tres capas distintas en el ala, formando un patrón entrecruzado cuando se superponen entre sí. Se desconoce la función de las actinofibrillas, al igual que el material exacto del que fueron hechas. Dependiendo de su composición exacta (queratina, músculo, estructuras elásticas, etc.), pueden haber sido agentes endurecedores o fortalecedores en la parte exterior del ala. [47]Las membranas de las alas también contenían una capa delgada de músculo, tejido fibroso y un sistema circulatorio complejo y único de vasos sanguíneos en bucle. [30] La combinación de actinofibrillas y capas musculares puede haber permitido al animal ajustar la flacidez y la comba de las alas . [45]
Como lo muestran las cavidades en los huesos de las alas de especies más grandes y el tejido blando conservado en al menos un espécimen, algunos pterosaurios extendieron su sistema de sacos de aire respiratorio hacia la membrana del ala. [48]
Partes del ala
La membrana del ala del pterosaurio se divide en tres unidades básicas. [49] El primero, llamado propatagium ("membrana delantera"), era la parte más adelantada del ala y estaba unida entre la muñeca y el hombro, creando el "borde de ataque" durante el vuelo. El braquiopatagio ("membrana del brazo") era el componente principal del ala, que se extendía desde el angosto dedo anular de la mano hasta las patas traseras. Finalmente, al menos algunos grupos de pterosaurios tenían una membrana que se extendía entre las piernas, posiblemente conectando o incorporando la cola, llamada uropatagium ; [49] la extensión de esta membrana no es segura, como estudios sobre Sordesparece sugerir que simplemente conectaba las patas pero no involucraba la cola (lo que la convierte en un cruropatagio ). Una interpretación común es que los pterosaurios no pterodactiloides tenían un uro / crurotagio más amplio estirado entre el largo quinto dedo del pie, y los pterodactiloides carecen de esos dedos y solo tienen membranas a lo largo de las piernas. [50]
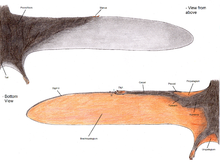
Ha habido una discusión considerable entre los paleontólogos sobre si las membranas principales del ala (braquiopatagia) se adhieren a las extremidades posteriores y, de ser así, dónde. Los fósiles del ramphorhynchoid Sordes , [51] el anurognathid Jeholopterus , [52] y un pterodactyloid de la Formación Santana parecen demostrar que la membrana del ala se adhirió a las extremidades posteriores, al menos en algunas especies. [53] Sin embargo, los murciélagos modernos y las ardillas voladorasmuestran una variación considerable en la extensión de las membranas de sus alas y es posible que, al igual que estos grupos, diferentes especies de pterosaurios tuvieran diferentes diseños de alas. De hecho, el análisis de las proporciones de las extremidades de los pterosaurios muestra que hubo una variación considerable, posiblemente reflejando una variedad de planos de alas. [54]
Los elementos óseos del brazo formaron un mecanismo para sostener y extender el ala. Cerca del cuerpo, el húmero o hueso de la parte superior del brazo es corto pero de constitución poderosa. [55] Tiene una gran cresta deltopectoral, a la que se unen los principales músculos de vuelo. [55] A pesar de las considerables fuerzas ejercidas sobre él, el húmero está hueco o neumatizado por dentro, reforzado por puntales óseos. [43] Los huesos largos del antebrazo, el cúbito y el radio , son mucho más largos que el húmero. [56] Probablemente eran incapaces de pronación .
Un hueso exclusivo de los pterosaurios, [57] conocido como pteroide, se conectaba a la muñeca y ayudaba a sostener la membrana delantera (el propatagio) entre la muñeca y el hombro. La evidencia de membranas entre los tres dedos libres de la extremidad anterior del pterosaurio sugiere que esta membrana delantera puede haber sido más extensa que la simple conexión del pteroide al hombro tradicionalmente representada en las restauraciones de vida. [30] La posición del hueso pteroide en sí ha sido controvertida. Algunos científicos, en particular Matthew Wilkinson, han argumentado que el pteroide apuntaba hacia adelante, extendiendo la membrana delantera y permitiéndole funcionar como un colgajo ajustable . [58]Este punto de vista fue contradicho en un artículo de 2007 de Chris Bennett, quien mostró que el pteroide no se articulaba como se pensaba anteriormente y no podía apuntar hacia adelante, sino que estaba dirigido hacia adentro, hacia el cuerpo, como se interpreta tradicionalmente. [59] Los especímenes de Changchengopterus pani y Darwinopterus linglongtaensis muestran el pteroide en articulación con el sincarpiano proximal, lo que sugiere que el pteroide se articulaba con la "silla" del radiale (sincarpiano proximal) y que tanto el pteroide como el carpiano preaxial eran centralia migrada. [60] [61]
La muñeca del pterosaurio consta de dos carpos (huesos de la muñeca) internos (proximales, al lado de los huesos largos del brazo) y cuatro externos (distal, al costado de la mano), excluyendo el hueso pteroide, que en sí mismo puede ser un carpo distal modificado. Los carpos proximales se fusionan en un "sincarpiano" en muestras maduras, mientras que tres de los carpos distales se fusionan para formar un sincarpiano distal. El carpiano distal restante, denominado aquí carpiano medial, pero que también se ha denominado carpiano lateral distal o preaxial, se articula en una faceta biconvexa alargada verticalmente en la superficie anterior del sincarpiano distal. El carpo medial tiene una fóvea cóncava profunda que se abre en sentido anterior, ventral y algo medial, dentro de la cual se articula el pteroide, según Wilkinson. [62]
En los pterodactiloides derivados como los pteranodoncianos y los azdarcoides , los metacarpianos I-III son pequeños y no se conectan al carpo, sino que cuelgan en contacto con el cuarto metacarpiano. [63] Con estas especies derivadas, el cuarto metacarpiano se ha alargado enormemente, por lo general igualando o excediendo la longitud de los huesos largos del antebrazo. [64] El quinto metacarpiano se había perdido. [55] En todas las especies, el primer y tercer dedo son mucho más pequeños que el cuarto, el "dedo ala", y contienen dos, tres y cuatro falanges respectivamente. [63] Los dedos más pequeños tienen garras, y el tamaño ungual varía entre especies. En nictosáuridoslos dedos de las extremidades anteriores además del dedo del ala se han perdido por completo. El dedo del ala representa aproximadamente la mitad o más de la longitud total del ala. [63] Normalmente consta de cuatro falanges. Sus longitudes relativas tienden a variar entre especies, lo que a menudo se ha utilizado para distinguir formas relacionadas. [63] La cuarta falange suele ser la más corta. Carece de garra y los nictosáuridos la han perdido por completo. Tiene una curva hacia atrás, lo que da como resultado una punta de ala redondeada, que reduce la resistencia inducida . El dedo del ala también está algo doblado hacia abajo. [64]
Al estar de pie, los pterosaurios probablemente descansaban sobre sus metacarpianos, con el ala exterior doblada hacia atrás. En esta posición, los lados "anteriores" de los metacarpianos se rotaron hacia atrás. Esto señalaría los dedos más pequeños oblicuamente hacia atrás. Según Bennett, esto implicaría que el dedo del ala, capaz de describir el arco más grande de cualquier elemento del ala, hasta 175 °, no se dobló por flexión sino por una extensión extrema. El ala se plegaba automáticamente cuando se doblaba el codo. [37] [65]
Pelvis
La pelvis de los pterosaurios era de tamaño moderado en comparación con el cuerpo en su conjunto. A menudo, los tres huesos pélvicos se fusionaron. [64] El hueso ilíaco era largo y bajo, sus hojas delantera y trasera se proyectaban horizontalmente más allá de los bordes de los huesos pélvicos inferiores. A pesar de esta longitud, la forma en forma de varilla de estos procesos indica que los músculos de las patas traseras unidos a ellos tenían una fuerza limitada. [37] En vista lateral, el hueso púbico estrecho se fusionó con el isquion ancho en una lámina isquiopúbica. A veces, las hojas de ambos lados también se fusionaron, cerrando la pelvis desde abajo y formando el canal pélvico. La articulación de la caderano estaba perforado y permitía una movilidad considerable a la pierna. [63] Se dirigió oblicuamente hacia arriba, impidiendo una posición perfectamente vertical de la pierna. [64]
El frente de los huesos púbicos articulado con una estructura única, los huesos prepúbicos emparejados. Juntos formaron una cúspide que cubría la parte posterior del vientre, entre la pelvis y las costillas del vientre. La movilidad vertical de este elemento sugiere una función en la respiración, compensando la relativa rigidez de la cavidad torácica. [63]
Extremidades traseras
Las patas traseras de los pterosaurios estaban fuertemente construidas, pero en relación con la envergadura de sus alas eran más pequeñas que las de las aves. Eran largos en comparación con la longitud del torso. [66] El fémur era bastante recto, con la cabeza formando solo un pequeño ángulo con el eje. [63] Esto implica que las piernas no se sostenían verticalmente debajo del cuerpo, sino que estaban algo extendidas. [66] La tibia se fusionaba a menudo con la parte superior del tobillo en un tibiotarsus que era más largo que el fémur. [66] Podría alcanzar una posición vertical al caminar. [66]El hueso de la pantorrilla tendía a ser delgado, especialmente en su extremo inferior que en formas avanzadas no llegaba al tobillo, a veces reduciendo la longitud total a un tercio. Por lo general, se fusionó con la espinilla. [63] El tobillo era una bisagra simple, "mesotarsal". [66] El metatarso , bastante largo y delgado, [67] siempre estaba extendido hasta cierto punto. [68] El pie era plantígrado, lo que significa que durante el ciclo de la marcha la planta del metatarso se presionaba contra el suelo. [67]
Hubo una clara diferencia entre los pterosaurios tempranos y las especies avanzadas con respecto a la forma del quinto dígito. Originalmente, el quinto metatarsiano era robusto y no muy acortado. Estaba conectado al tobillo en una posición más alta que los otros metatarsianos. [67] Presentaba un quinto dedo del pie largo, ya menudo curvado, móvil sin garras que constaba de dos falanges. [68] La función de este elemento ha sido enigmática. Se pensaba que los animales dormían boca abajo como murciélagos, colgados de las ramas y usando el quinto dedo del pie como ganchos. Otra hipótesis sostenía que estiraban la braquiopatagia, pero en los fósiles articulados el quinto dedo siempre está flexionado hacia la cola. [67]Más tarde se hizo popular suponer que estos dedos extendían un uropatagium o cruropatagium entre ellos. Como los quintos dedos estaban en la parte externa de los pies, tal configuración solo habría sido posible si estos giraran sus frentes hacia afuera en vuelo. [67] Tal rotación podría ser causada por una abducción del fémur, lo que significa que las piernas estarían abiertas. Esto también convertiría los pies en una posición vertical. [67] Entonces podrían actuar como timones para controlar la guiñada. Algunas muestras muestran membranas entre los dedos de los pies, [69]permitiéndoles funcionar como superficies de control de vuelo. El uropatagium o cruropatagium controlaría el tono. Al caminar, los dedos de los pies pueden flexionarse hacia arriba para levantar la membrana del suelo. En Pterodactyloidea, el quinto metatarsiano estaba muy reducido y el quinto dedo, si estaba presente, poco más que un muñón. [70] Esto sugiere que sus membranas se dividieron, aumentando la maniobrabilidad de vuelo. [50]
Los dedos del primero al cuarto eran largos. Tenían dos, tres, cuatro y cinco falanges respectivamente. [66] A menudo, el tercer dedo era el más largo; a veces el cuarto. Las articulaciones planas indican una movilidad limitada. Estos dedos tenían garras, pero las garras eran más pequeñas que las garras de las manos. [68]
Tejidos blandos
Las raras condiciones que permitieron la fosilización de restos de pterosaurios, a veces también preservaron tejidos blandos. La fotografía moderna sincrotrón o con luz ultravioleta ha revelado muchos rastros que no son visibles a simple vista. [71] A menudo se les llama imprecisamente "impresiones", pero en su mayoría consisten en petrificaciones , moldes naturales y transformaciones del material original. Pueden incluir crestas de cuerno, picos o vainas de garras, así como las diversas membranas de vuelo. Excepcionalmente, se conservaron los músculos. [72] Los parches cutáneos muestran pequeñas escamas redondas que no se superponen en las plantas de los pies, los tobillos y los extremos de los metacarpianos. [73]Cubrieron almohadillas que amortiguan el impacto de caminar. Se desconocen las escamas de otras partes del cuerpo. [74]
Pycnofibers
La mayoría o todos los pterosaurios tenían filamentos similares a pelos conocidos como picnofibras en la cabeza y el torso. [75] El término "picnofibra", que significa "filamento denso", fue acuñado por el paleontólogo Alexander Kellner y sus colegas en 2009. [47] Las picnofibras eran estructuras únicas similares, pero no homólogas (que comparten un origen común) con el pelo de los mamíferos , un ejemplo de evolución convergente . [51] Un tegumento difuso fue informado por primera vez de un espécimen de Scaphognathus crassirostris en 1831 por Georg Augustus Goldfuss , [76]pero se había dudado mucho. Desde la década de 1990, los hallazgos de pterosaurios y el examen histológico y ultravioleta de especímenes de pterosaurios han proporcionado una prueba incontrovertible: los pterosaurios tenían capas de picnofibras. Sordes pilosus (que se traduce como "demonio peludo") y Jeholopterus ninchengensis muestran picnofibras en la cabeza y el cuerpo.
La presencia de picnofibras indica claramente que los pterosaurios eran endotérmicos (de sangre caliente). Ayudaron a la termorregulación, como es común en los animales de sangre caliente que necesitan aislamiento para evitar una pérdida excesiva de calor. [75] Las picnofibras eran filamentos cortos y flexibles, de entre cinco y siete milímetros de largo y una estructura bastante simple con un canal central hueco. [75] Las pieles de pterosaurio podrían haber sido comparables en densidad a la de muchos mamíferos mesozoicos. [b] [75]
Relación con las plumas
Los filamentos de pterosaurio podrían compartir un origen común con las plumas, como especularon en 2002 Czerkas y Ji. [31] En 2009, Kellner concluyó que las picnofibras estaban estructuradas de manera similar a las protoplumas de terópodos . [47] Otros no estaban convencidos, considerando la diferencia con las "púas" encontradas en muchos de los especímenes de maniraptoran con forma de pájaro demasiado fundamental. [75]
Un estudio de 2018 de los restos de dos pequeños pterosaurios de la edad jurásica de Mongolia Interior , China , encontró que los pterosaurios tenían una amplia gama de formas y estructuras de picnofibras, a diferencia de las estructuras homogéneas que generalmente se suponía que los cubrían. Algunos de estos tenían extremos deshilachados, muy similares en estructura a cuatro tipos de plumas diferentes conocidos de aves u otros dinosaurios, pero casi nunca conocidos de pterosaurios antes del estudio, lo que sugiere homología. [77] [78] Se publicó una respuesta a este estudio en 2020, donde se sugirió que las estructuras que se ven en los anurognátidosfueron en realidad el resultado de la descomposición de las aktinofibrillas: un tipo de fibra que se usa para fortalecer y endurecer el ala. [79] Sin embargo, en respuesta a esto, los autores del artículo de 2018 señalan el hecho de que la presencia de las estructuras se extiende más allá del patagium y la presencia de aktinofibrillas y filamentos en Jeholopterus ningchengensis [80] y Sordes pilosus . [81] Las diversas formas de estructura de filamentos presentes en los anurognátidos en el estudio de 2018 también requerirían una forma de descomposición que causaría las diferentes formas de 'filamentos' observadas. Por tanto, concluyen que la interpretación más parsimoniosa de las estructuras es que son proto-plumas filamentosas.[82]
Historia del descubrimiento
Primero encuentra
Los fósiles de pterosaurios son muy raros debido a su estructura ósea ligera. Los esqueletos completos generalmente solo se pueden encontrar en capas geológicas con condiciones de conservación excepcionales, las llamadas Lagerstätten . Las piezas de una de esas Lagerstätte , la piedra caliza Solnhofen del Jurásico tardío en Baviera , [83] fueron muy buscadas por coleccionistas ricos. [84] En 1784, el naturalista italiano Cosimo Alessandro Collini fue el primer científico de la historia en describir un fósil de pterosaurio. [85]En ese momento, los conceptos de evolución y extinción solo se desarrollaron de manera imperfecta. Por lo tanto, la extraña construcción del pterosaurio fue impactante, ya que no podía asignarse claramente a ningún grupo de animales existente. [86] El descubrimiento de los pterosaurios jugaría un papel importante en el progreso de la paleontología y la geología modernas. [87] Si tales criaturas todavía estuvieran vivas, solo el mar era un hábitat creíble y Collini sugirió que podría ser un animal nadador que usaba sus largas extremidades delanteras como remos. [88] Algunos científicos continuaron apoyando la interpretación acuática incluso hasta 1830, cuando el zoólogo alemán Johann Georg Wagler sugirió que Pterodactylus usaba sus alas como aletas y estaba afiliado conIctiosaurios y plesiosauria . [89]
En 1800, Johann Hermann sugirió por primera vez que representaba una criatura voladora en una carta a Georges Cuvier . Cuvier estuvo de acuerdo en 1801, entendiendo que era un reptil volador extinto. [90] En 1809, acuñó el nombre de Ptéro-Dactyle , "dedo de ala". [91] Esto fue latinizado en 1815 a Pterodactylus . [92] Al principio, la mayoría de las especies se asignaron a este género y, finalmente, "pterodáctilo" se aplicó popular e incorrectamente a todos los miembros de Pterosauria. [15] Hoy en día, los paleontólogos limitan el término al género Pterodactylus o miembros de Pterodactyloidea . [dieciséis]
En 1812 y 1817, Samuel Thomas von Soemmerring redescribió el espécimen original y uno adicional. [93] Los vio como afiliados a pájaros y murciélagos. Aunque estaba equivocado en esto, su "modelo de murciélago" sería muy influyente durante el siglo XIX. [94] En 1843, Edward Newman pensó que los pterosaurios eran marsupiales voladores . [95] Como el modelo de murciélago describía correctamente a los pterosaurios como peludos y de sangre caliente, se acercaba mejor a la fisiología verdadera de los pterosaurios que el "modelo reptil" de Cuvier. En 1834, Johann Jakob Kaup acuñó el término Pterosauria. [96]
Ampliando la investigación
En 1828, Mary Anning en Inglaterra encontró el primer género de pterosaurios fuera de Alemania, [97] por Richard Owen llamado Dimorphodon , también el primer pterosaurio no pterodactiloide conocido. [98] Más adelante en el siglo, Cambridge Greensand del Cretácico Temprano produjo miles de fósiles de pterosaurios, que sin embargo, eran de mala calidad, y consistían principalmente en fragmentos fuertemente erosionados. [99] Sobre la base de estos, sin embargo, se nombrarían numerosos géneros y especies. [87] Muchos fueron descritos por Harry Govier Seeley , en ese momento el principal experto inglés en el tema, quien también escribió el primer libro de pterosaurios,Ornithosauria , [100] y en 1901 el primer libro popular, [87] Dragons of the Air . Seeley pensó que los pterosaurios eran criaturas dinámicas y de sangre caliente, estrechamente relacionadas con las aves. [101] Anteriormente, el evolucionista St. George Jackson Mivart había sugerido que los pterosaurios eran los antepasados directos de las aves. [102] Owen se opuso a las opiniones de ambos hombres, viendo a los pterosaurios como reptiles "verdaderos" de sangre fría. [103]
En los Estados Unidos, Othniel Charles Marsh en 1870 descubrió Pteranodon en Niobrara Chalk , entonces el pterosaurio más grande conocido, [103] el primer desdentado y el primero de América. [104] Estas capas también produjeron miles de fósiles, [104] también incluyendo esqueletos relativamente completos que fueron preservados en tres dimensiones en lugar de estar fuertemente comprimidos como con los especímenes de Solnhofen. Esto condujo a una mejor comprensión de muchos detalles anatómicos, [104] como la naturaleza hueca de los huesos.
Mientras tanto, los hallazgos del Solnhofen habían continuado, lo que representa la mayoría de los especímenes completos de alta calidad descubiertos. [105] Permitieron identificar la mayoría de los nuevos taxones basales, como Rhamphorhynchus , Scaphognathus y Dorygnathus . [105] Este material dio origen a una escuela alemana de investigación de pterosaurios, que consideraba a los reptiles voladores como las contrapartes mesozoicas activas, peludas y de sangre caliente de los murciélagos y pájaros modernos. [106] En 1882, Marsh y Karl Alfred Zittel publicaron estudios sobre las membranas de las alas de especímenes de Rhamphorhynchus . [107] [108]Los estudios alemanes continuaron hasta bien entrada la década de 1930, describiendo nuevas especies como Anurognathus . En 1927, Ferdinand Broili descubrió folículos pilosos en la piel de los pterosaurios, [109] y la paleoneuróloga Tilly Edinger determinó que los cerebros de los pterosaurios se parecían más a los de las aves que a los de los modernos reptiles de sangre fría. [110]
Por el contrario, los paleontólogos ingleses y estadounidenses a mediados del siglo XX perdieron en gran medida el interés por los pterosaurios. Los vieron como experimentos evolutivos fallidos, de sangre fría y escamosos, que apenas podían volar, las especies más grandes solo podían deslizarse, siendo obligadas a trepar árboles o arrojarse desde acantilados para lograr un despegue. En 1914, por primera vez, la aerodinámica de los pterosaurios fue analizada cuantitativamente por Ernest Hanbury Hankin y David Meredith Seares Watson , pero interpretaron al Pteranodon como un planeador puro. [111] Se realizaron pocas investigaciones sobre el grupo durante las décadas de 1940 y 1950. [87]
Renacimiento de pterosaurio
La situación de los dinosaurios era comparable. A partir de la década de 1960, se produjo un renacimiento de los dinosaurios , un rápido aumento en el número de estudios e ideas críticas, influenciado por el descubrimiento de fósiles adicionales de Deinonychus , cuyos rasgos espectaculares refutaban lo que se había convertido en una ortodoxia arraigada. En 1970, igualmente la descripción del pterosaurio peludo Sordes inició lo que Robert Bakker denominó un renacimiento de los pterosaurios. [112] Especialmente Kevin Padian propagó los nuevos puntos de vista, publicando una serie de estudios que mostraban a los pterosaurios como animales de sangre caliente, activos y corriendo. [113] [114] [115]Esto coincidió con un renacimiento de la escuela alemana a través del trabajo de Peter Wellnhofer , quien en la década de 1970 sentó las bases de la ciencia moderna de los pterosaurios. [83] En 1978, publicó el primer libro de texto de pterosaurios, [116] el Handbuch der Paläoherptologie, Teil 19: Pterosauria , [117] y en 1991 el segundo libro de ciencia popular sobre pterosaurios, [116] la Encyclopedia of Pterosaurs . [118]
Este desarrollo se aceleró gracias a la explotación de dos nuevos Lagerstätten . [116] Durante la década de 1970, la Formación Santana del Cretácico Temprano en Brasil comenzó a producir nódulos de cretáceo que, aunque a menudo de tamaño limitado y la integridad de los fósiles que contenían, partes esqueléticas tridimensionales de pterosaurio perfectamente conservadas. [116] Los institutos alemanes y holandeses compraron tales nódulos de cazadores furtivos de fósiles y los prepararon en Europa, lo que permitió a sus científicos describir muchas especies nuevas y revelar una fauna completamente nueva. Pronto, investigadores brasileños, entre ellos Alexander Kellner , interceptaron el comercio y nombraron aún más especies.
Aún más productiva fue la Biota Jehol china del Cretácico Temprano de Liaoning que desde la década de 1990 ha producido cientos de fósiles bidimensionales exquisitamente conservados, que a menudo muestran restos de tejidos blandos. Investigadores chinos como Lü Junchang han vuelto a nombrar muchos taxones nuevos. A medida que los descubrimientos también aumentaron en otras partes del mundo, se produjo un aumento repentino en el total de géneros nombrados. En 2009, cuando habían aumentado a aproximadamente noventa, este crecimiento no mostró signos de estabilización. [119] En 2013, MP Witton indicó que el número de especies de pterosaurios descubiertas había aumentado a 130. [120]Más del noventa por ciento de los taxones conocidos han sido nombrados durante el "renacimiento". Muchos de ellos pertenecían a grupos cuya existencia se desconocía. [116] Los avances en la potencia informática permitieron determinar sus complejas relaciones mediante el método cuantitativo de la cladística . Los fósiles nuevos y antiguos arrojaron mucha más información cuando se sometieron a la luz ultravioleta moderna o fotografías de roentgen, o escaneos CAT . [121] Se aplicaron conocimientos de otros campos de la biología a los datos obtenidos. [121] Todo esto dio como resultado un progreso sustancial en la investigación de pterosaurios, lo que hizo que los relatos más antiguos de los libros de divulgación científica quedaran completamente obsoletos.
Evolución y extinción
Orígenes
Debido a que la anatomía de los pterosaurios se ha modificado mucho para el vuelo, y hasta ahora no se han descrito predecesores fósiles de transición inmediatos , la ascendencia de los pterosaurios no se comprende completamente. [122] Los pterosaurios más antiguos conocidos ya estaban completamente adaptados a un estilo de vida volador. Desde Seeley, se reconoció que era probable que los pterosaurios tuvieran su origen en los "arcosaurios", lo que hoy se llamaría Archosauromorpha . En la década de 1980, los primeros análisis cladísticos encontraron que eran avemetatarsianos (arcosaurios más cercanos a los dinosaurios que a los cocodrilos).). Como esto los convertiría también en parientes bastante cercanos de los dinosaurios, Kevin Padian consideró que estos resultados confirmaban su interpretación de los pterosaurios como animales bípedos de sangre caliente. Debido a que estos primeros análisis se basaron en un número limitado de taxones y caracteres, sus resultados fueron intrínsecamente inciertos. Varios investigadores influyentes que rechazaron las conclusiones de Padian ofrecieron hipótesis alternativas. David Unwin propuso una ascendencia entre los Archosauromorpha basales, específicamente formas de cuello largo (" protorosaurios ") como los tanystropheids . También se sugirió una ubicación entre arcosauriformes basales como Euparkeria . [23]Algunos arcosauromorfos basales parecen a primera vista ser buenos candidatos para parientes cercanos de pterosaurios debido a su anatomía de extremidades largas; un ejemplo es Sharovipteryx , un "protorosaurio" con membranas de piel en sus patas traseras que probablemente se usa para deslizarse. [123] Un estudio de 1999 de Michael Benton encontró que los pterosaurios eran avemetatarsalianos estrechamente relacionados con Scleromochlus , y denominó al grupo Ornithodira para abarcar pterosaurios y dinosaurios . [124]
Dos investigadores, S. Christopher Bennett en 1996, [125] y el paleoartista David Peters en 2000, publicaron análisis que encontraron que los pterosaurios eran protorosaurios o estaban estrechamente relacionados con ellos. Sin embargo, Peters recopiló datos anatómicos novedosos utilizando una técnica no verificada llamada "Segregación de gráficos digitales" (DGS), que implica el seguimiento digital de imágenes de fósiles de pterosaurios utilizando un software de edición de fotografías. [126] Bennett solo recuperó a los pterosaurios como parientes cercanos de los protorosaurios después de eliminar las características de las patas traseras de su análisis, para probar la posibilidad de una evolución convergente basada en la locomoción entre pterosaurios y dinosaurios.. Una respuesta de 2007 de Dave Hone y Michael Benton no pudo reproducir este resultado, encontrando que los pterosaurios están estrechamente relacionados con los dinosaurios incluso sin los personajes de las patas traseras. También criticaron a David Peters por sacar conclusiones sin acceso a la evidencia primaria, es decir, los propios fósiles de pterosaurios. [127] Hone y Benton concluyeron que, aunque se necesitan más pterosauromorfos basales para aclarar sus relaciones, la evidencia actual indica que los pterosaurios son avemetatarsalianos, ya sea como el grupo hermano de Scleromochlus o como una rama entre este último y Lagosuchus . [127] Un análisis filogenético centrado en arcosaurios de 2011 realizado por Sterling Nesbittse benefició de muchos más datos y encontró un fuerte apoyo para que los pterosaurios fueran avemetatarsianos, aunque Scleromochlus no se incluyó debido a su mala conservación. [128] Un estudio centrado en arcosauromorfos de 2016 realizado por Martin Ezcurra incluyó varios parientes de pterosaurios propuestos, pero también encontró que los pterosaurios estaban más cerca de los dinosaurios y no estaban relacionados con taxones más basales. [129] A partir de su análisis de 1996, Bennett publicó un estudio de 2020 sobre Scleromochlus que argumentó que tanto Scleromochlus como los pterosaurios eran arcosauromorfos no arcosaurios, aunque no particularmente relacionados entre sí. [130] Por el contrario, un estudio posterior de 2020 propuso que lagerpetid los arcosaurios eran el clado hermano de los pterosaurios. [131] Esto se basó en cráneos fósiles y extremidades anteriores recientemente descritos que muestran varias similitudes anatómicas con pterosaurios y reconstrucciones de cerebros lagerpétidos y sistemas sensoriales basados en tomografías computarizadas que también muestran similitudes neuroanatómicas con pterosaurios. [132] [133]
Un problema relacionado es el origen del vuelo de los pterosaurios. [134] Al igual que con las aves, las hipótesis se pueden ordenar en dos variedades principales: "molido" o "árbol abajo". Trepar a un árbol causaría que la altura y la gravedad proporcionaran tanto la energía como una fuerte presión de selección para un vuelo incipiente. [ aclaración necesaria ] Rupert Wild en 1983 propuso un hipotético "propterosaurus": un animal arbóreo parecido a un lagarto que desarrolla una membrana entre sus extremidades, primero para lanzarse en paracaídas con seguridad y luego, alargando gradualmente el cuarto dedo, para deslizarse. [135]Sin embargo, los resultados cladísticos posteriores no se ajustaron bien a este modelo. Ni los protorosaurios ni los ornitodiranos son biológicamente equivalentes a los lagartos. Además, la transición entre vuelo sin motor y con aleteo no se comprende bien. Estudios más recientes sobre la morfología de las patas traseras de los pterosaurios basales parecen reivindicar una conexión con Scleromochlus . Al igual que este arcosaurio, los linajes de pterosaurios basales tienen patas traseras plantígradas que muestran adaptaciones para la saltación. [136]
Extinción
Alguna vez se pensó que la competencia con las especies de aves tempranas podría haber resultado en la extinción de muchos de los pterosaurios. [137] Parte de esto se debe al hecho de que se solía pensar que a fines del Cretácico, solo estaban presentes grandes especies de pterosaurios (ya no es cierto; ver más abajo). Se pensaba que las especies más pequeñas se habían extinguido y que su nicho estaba lleno de aves. [138] Sin embargo, la disminución de pterosaurios (si realmente está presente) parece no estar relacionada con la diversidad de aves, ya que la superposición ecológica entre los dos grupos parece ser mínima. [139] De hecho, al menos algunos nichos de aves fueron reclamados por pterosaurios antes del evento KT . [140]Al final del período Cretácico, el evento de extinción Cretácico-Paleógeno , que acabó con todos los dinosaurios no aviares y también la mayoría de los dinosaurios aviares, y muchos otros animales, parece haber tomado también a los pterosaurios.
A principios de la década de 2010, se descubrieron varios taxones nuevos de pterosaurios que datan del Campaniano / Maastrichtiano, como los ornitoqueíridos Piksi y " Ornithocheirus ", posibles pteranodóntidos y nictosáuridos , varios tapejaridos y el indeterminado Navajodactylus no azdárquico . [141] [142] Los pequeños pterosaurios azdarcoides también estaban presentes en el Campaniano. Esto sugiere que las faunas de pterosaurios del Cretácico tardío eran mucho más diversas de lo que se pensaba anteriormente, posiblemente ni siquiera habiendo disminuido significativamente desde el Cretácico temprano.
Aparentemente, especies de pterosaurios de pequeño tamaño estaban presentes en la Formación Csehbánya , lo que indica una mayor diversidad de pterosaurios del Cretácico Superior de lo que se contaba anteriormente. [143] Los hallazgos recientes de un azdárquido adulto del tamaño de un gato pequeño indican además que los pterosaurios pequeños del Cretácico tardío en realidad podrían haberse conservado en raras ocasiones en el registro fósil, ayudado por el hecho de que existe un fuerte sesgo contra los vertebrados terrestres de pequeño tamaño. como los dinosaurios juveniles , y que su diversidad en realidad podría haber sido mucho mayor de lo que se pensaba anteriormente. [144]
Al menos algunos pterosaurios no pterodactiloides sobrevivieron hasta el Cretácico tardío , postulando una situación de taxones de Lázaro para las faunas de pterosaurios del Cretácico tardío. [145]
Clasificación y filogenia
En taxonomía filogenética , el clado Pterosauria generalmente se ha definido como basado en nodos y anclado a varios taxones ampliamente estudiados, así como a aquellos que se cree que son primitivos. Un estudio de 2003 definió a Pterosauria como "El ancestro común más reciente de Anurognathidae, Preondactylus y Quetzalcoatlus y todos sus descendientes". [146] Sin embargo, este tipo de definición dejaría inevitablemente a cualquier especie relacionada que sea un poco más primitiva fuera de la Pterosauria. Para remediar esto, se propuso una nueva definición que anclaría el nombre no a ninguna especie en particular sino a una característica anatómica, la presencia de un cuarto dedo agrandado que sostiene la membrana del ala. [147]Esta definición "basada en apomorofia" fue adoptada por el PhyloCode en 2020 como "[E] l clado caracterizado por la apomorfia del cuarto dedo manual hipertrofiado para soportar una membrana del ala, heredada por Pterodactylus (originalmente Ornithocephalus ) antiquus (Sömmerring 1812)". [148] Un clado más amplio, Pterosauromorpha , ha sido definido como todos los ornithodirans más estrechamente relacionados con los pterosaurios que con los dinosaurios . [149]
Históricamente, la clasificación interna de los pterosaurios ha sido difícil, porque había muchas lagunas en el registro fósil . A partir del siglo XXI, nuevos descubrimientos están llenando estos vacíos y dando una mejor imagen de la evolución de los pterosaurios. Tradicionalmente, estaban organizados en dos subórdenes : los Rhamphorhynchoidea , un grupo "primitivo" de pterosaurios de cola larga, y los Pterodactyloidea , pterosaurios "avanzados" con colas cortas. [23] Sin embargo, esta división tradicional se ha abandonado en gran medida. Rhamphorhynchoidea es un parafilético(antinatural), ya que los pterodactiloides evolucionaron directamente de ellos y no de un ancestro común, por lo que, con el uso creciente de cladística , ha caído en desgracia entre la mayoría de los científicos. [120] [150]
Las relaciones precisas entre los pterosaurios aún no se han resuelto. Muchos estudios sobre las relaciones de los pterosaurios en el pasado han incluido datos limitados y fueron muy contradictorios. Sin embargo, los estudios más recientes que utilizan conjuntos de datos más grandes están comenzando a aclarar las cosas. El cladograma (árbol genealógico) a continuación sigue un análisis filogenético presentado por Longrich, Martill y Andres en 2018, con nombres de clado después de Andres et al (2014). [140] [1]
| Pterosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiología
Vuelo
La mecánica del vuelo de los pterosaurios no se comprende ni se modela completamente en este momento. [151] [152]
Katsufumi Sato, un científico japonés, hizo cálculos utilizando aves modernas y concluyó que era imposible que un pterosaurio se mantuviera en el aire. [151] En el libro Postura, locomoción y paleoecología de los pterosaurios se teoriza que pudieron volar debido a la atmósfera densa y rica en oxígeno del período Cretácico tardío. [153] Sin embargo, tanto Sato como los autores de Posture, Locomotion, and Paleoecology of Pterosaurs basaron su investigación en las ahora obsoletas teorías de que los pterosaurios son parecidos a las aves marinas, y el límite de tamaño no se aplica a los pterosaurios terrestres, como los azdárquidos y tapejarids . Además, Darren Naishconcluyó que las diferencias atmosféricas entre el presente y el Mesozoico no eran necesarias para el tamaño gigante de los pterosaurios. [154]
Otro tema que ha sido difícil de entender es cómo despegaron . Las sugerencias anteriores eran que los pterosaurios eran en gran parte animales deslizantes de sangre fría, que derivaban el calor del medio ambiente como los lagartos modernos, en lugar de quemar calorías. En este caso, no estaba claro cómo los más grandes de enorme tamaño, con un ineficiente metabolismo de sangre fría, podían manejar una estrategia de despegue similar a la de un pájaro, usando solo las extremidades traseras para generar empuje para volar. Investigaciones posteriores muestran que, en cambio, son de sangre caliente y tienen poderosos músculos de vuelo, y usan los músculos de vuelo para caminar como cuadrúpedos. [155] Mark Witton de la Universidad de Portsmouth y Mike Habib de la Universidad Johns Hopkinssugirió que los pterosaurios usaban un mecanismo de salto para obtener vuelo. [156] El tremendo poder de sus patas delanteras aladas les permitiría despegar con facilidad. [155] Una vez en el aire, los pterosaurios podrían alcanzar velocidades de hasta 120 km / h (75 mph) y viajar miles de kilómetros. [156]
En 1985, la Institución Smithsonian encargó al ingeniero aeronáutico Paul MacCready que construyera un modelo de trabajo a media escala de Quetzalcoatlus northropi . La réplica se lanzó con un cabrestante terrestre. Voló varias veces en 1986 y fue filmado como parte de la película IMAX del Smithsonian On the Wing . [157] [158]
Sacos de aire y respiración
Un estudio de 2009 mostró que los pterosaurios tenían un sistema de pulmón y saco de aire y una bomba de respiración esquelética controlada con precisión, que respalda un modelo de ventilación pulmonar de flujo continuo en los pterosaurios, análogo al de las aves. La presencia de un sistema de saco de aire subcutáneo en al menos algunos pterodactiloides habría reducido aún más la densidad del animal vivo. [48] Como los cocodrilos modernos, los pterosaurios parecían haber tenido un pistón hepático , ya que sus cinturas escapulares y pectorales eran demasiado inflexibles para mover el esternón como en las aves, y poseían una fuerte gastralia . [159] Por lo tanto, su sistema respiratorio tenía características comparables a las de los dos clados de arcosaurios modernos.
Sistema nervioso
Un estudio de rayos X de las cavidades cerebrales de los pterosaurios reveló que los animales ( Rhamphorhynchus muensteri y Anhanguera santanae ) tenían flóculos masivos. El flóculo es una región del cerebro que integra señales de articulaciones, músculos, piel y órganos del equilibrio. [17] Los flóculos de los pterosaurios ocuparon el 7,5% de la masa cerebral total de los animales, más que en cualquier otro vertebrado. Las aves tienen flóculos inusualmente grandes en comparación con otros animales, pero estos solo ocupan entre el 1 y el 2% de la masa cerebral total. [17]
El flóculo envía señales neurales que producen pequeños movimientos automáticos en los músculos del ojo. Estos mantienen estable la imagen en la retina de un animal. Los pterosaurios pueden haber tenido un flóculo tan grande debido al gran tamaño de sus alas, lo que significaría que había mucha más información sensorial para procesar. [17] La baja masa relativa de los flóculos en las aves también se debe a que las aves tienen un cerebro mucho más grande en general; aunque esto se ha considerado una indicación de que los pterosaurios vivían en un entorno estructuralmente más simple o tenían un comportamiento menos complejo en comparación con las aves, [160] estudios recientes de cocodrilos y otros reptiles muestran que es común que los saurópsidos alcancen altos niveles de inteligencia con cerebros pequeños. [161] Estudios sobre el endocast deAllkaruen muestra que la evolución del cerebro en pterodactiloides fue un proceso modular. [162]
Movimiento de tierra

Las cuencas de las caderas de los pterosaurios están orientadas ligeramente hacia arriba, y la cabeza del fémur (hueso del muslo) está solo moderadamente hacia adentro, lo que sugiere que los pterosaurios tenían una postura erguida. Habría sido posible levantar el muslo a una posición horizontal durante el vuelo, como hacen las lagartijas planeadoras.
Hubo un debate considerable sobre si los pterosaurios deambulaban como cuadrúpedos o como bípedos . En la década de 1980, el paleontólogo Kevin Padian sugirió que los pterosaurios más pequeños con patas traseras más largas, como Dimorphodon , podrían haber caminado o incluso correr bípedo, además de volar, como los corredores de carretera . [115] Sin embargo, una gran cantidad de huellas de pterosaurios se encontraron más tarde con una pata trasera distintiva de cuatro dedos y una pata delantera de tres dedos; estas son las huellas inconfundibles de pterosaurios caminando a cuatro patas. [163] [164]
Las huellas fósiles muestran que los pterosaurios estaban con todo el pie en contacto con el suelo ( plantígrado ), de una manera similar a muchos mamíferos como los humanos y los osos . Huellas de azdárquidos y varias especies no identificadas muestran que los pterosaurios caminaban con una postura erguida con sus cuatro extremidades sostenidas casi verticalmente debajo del cuerpo, una postura de eficiencia energética utilizada por la mayoría de las aves y mamíferos modernos, en lugar de las extremidades extendidas de los reptiles modernos. [69] [155] De hecho, las extremidades erectas pueden ser omnipresentes en los pterosaurios. [136]

Aunque tradicionalmente se describe como torpe y torpe cuando está en el suelo, la anatomía de algunos pterosaurios (particularmente los pterodactiloides) sugiere que eran caminantes y corredores competentes. [165] Los primeros pterosaurios se han considerado durante mucho tiempo como locomotoras particularmente engorrosas debido a la presencia de una gran crurotagia , pero también parecen haber sido generalmente eficientes en el suelo. [136]
Los huesos de las extremidades anteriores de los azdárquidos y ornitoqueíridos eran inusualmente largos en comparación con otros pterosaurios y, en los azdárquidos, los huesos del brazo y la mano (metacarpianos) estaban particularmente alargados. Además, en su conjunto, las extremidades delanteras de los azdárquidos tenían proporciones similares a las de los mamíferos ungulados que corren rápidamente . Sus extremidades traseras, por otro lado, no estaban construidas para la velocidad, pero eran largas en comparación con la mayoría de los pterosaurios y permitían una gran longitud de zancada. Si bien los pterosaurios azdárquidos probablemente no podrían correr, habrían sido relativamente rápidos y energéticamente eficientes. [69]
El tamaño relativo de las manos y los pies de los pterosaurios (en comparación con los animales modernos como las aves) puede indicar el tipo de estilo de vida que llevan los pterosaurios en el suelo. Los pterosaurios azdárquidos tenían pies relativamente pequeños en comparación con el tamaño de su cuerpo y la longitud de las piernas, y la longitud de los pies era de sólo un 25-30% de la longitud de la parte inferior de la pierna. Esto sugiere que los azdárquidos se adaptaron mejor a caminar sobre terreno seco y relativamente sólido. Pteranodon tenía pies un poco más grandes (47% de la longitud de la tibia ), mientras que los pterosaurios que se alimentaban por filtración como los ctenochasmatoides tenían pies muy grandes (69% de la longitud tibial en Pterodactylus , 84% en Pterodaustro ), adaptados para caminar en suelos blandos y fangosos. similar a las aves zancudas modernas. [69]Aunque claramente son lanzadores de patas delanteras, los pterosaurios basales tienen patas traseras bien adaptadas para saltar, lo que sugiere una conexión con arcosaurios como Scleromochlus . [136]
Natación
Las huellas de los ctenochasmatoides indican que estos pterosaurios nadaban usando sus patas traseras. En general, estos tienen patas traseras grandes y torsos largos, lo que indica que probablemente estaban más adaptados para nadar que otros pterosaurios. [166] Por el contrario, los pteranodoncistas tienen varias especiaciones en el húmero que se interpretan como sugestivas de una versión acuática del típico lanzamiento cuadrúpedo, y varios boreopteridos similares debieron alimentarse mientras nadaban, ya que parecen incapaces de ceder como un ave fragata . [166] Estas adaptaciones también se observan en pterosaurios terrestres como los azdárquidos , que presumiblemente todavía necesitaban lanzarse desde el agua en caso de que se encontraran en ella. ElEl nyctosaurid Alcione puede mostrar adaptaciones para el buceo propulsado por alas, como los alcatraces y las aves tropicales modernas . [167]
Dieta y hábitos alimentarios
Tradicionalmente, casi todos los pterosaurios eran vistos como piscívoros que se alimentaban en la superficie o devoradores de peces, una visión que todavía domina la ciencia popular. Hoy en día, se cree que muchos grupos de pterosaurios eran carnívoros terrestres, omnívoros o insectívoros.
Desde el principio se reconoció que los pequeños Anurognathidae eran insectívoros aéreos y nocturnos. Con articulaciones muy flexibles en el dedo del ala, una forma de ala ancha y triangular, ojos grandes y cola corta, estos pterosaurios probablemente eran análogos a los chotacabras o murciélagos insectívoros existentes, siendo capaces de una alta maniobrabilidad a velocidades relativamente bajas. [168]
Las interpretaciones de los hábitos de los grupos basales han cambiado profundamente. Dimorphodon , concebido como un análogo del frailecillo en el pasado, se indica por la estructura de su mandíbula, su forma de andar y su escasa capacidad de vuelo, como un depredador terrestre / semiarbóreo de pequeños mamíferos, escamatos e insectos grandes. [169] Su dentición robusta hizo que Campylognathoides fuera visto como un generalista o un depredador terrestre de pequeños vertebrados, pero el húmero muy robusto y la morfología del ala de aspecto alto sugieren que pudo haber sido capaz de agarrar presas en el ala. [170] El pequeño Carniadactylus insectívoro y el Eudimorphodon más grandeEran animales muy aéreos y voladores rápidos y ágiles con alas largas y robustas. Se ha encontrado Eudimorphodon con restos de pescado en su estómago, pero su dentición sugiere una dieta oportunista. Austriadactylus y Caviramus de alas delgadas probablemente eran generalistas terrestres / semiarbóreos. Es probable que el caviramus tuviera una fuerte fuerza de mordida, lo que indica una adaptación hacia alimentos duros que podrían haber sido masticados en vista del desgaste de los dientes. [171]
Algunos Rhamphorhynchidae , como el propio Rhamphorhynchus o Dorygnathus , comían pescado con alas largas y delgadas, dentición en forma de aguja y mandíbulas largas y delgadas. Sericipterus , Scaphognathus y Harpactognathus tenían mandíbulas y dientes más robustos (que eran zifodontes, en forma de daga, en Sericipterus ) y alas más cortas y anchas. Estos eran depredadores terrestres / aéreos de vertebrados [172] o generalistas parecidos a los córvidos . [173] Wukongopteridae como Darwinopterusfueron considerados por primera vez depredadores aéreos. Al carecer de una estructura de mandíbula robusta o de poderosos músculos voladores, ahora se los considera insectívoros arbóreos o semiterrestres. Darwinopterus robustidens , en particular, parece haber sido un especialista en escarabajos. [174]
Entre los pterodactiloides, existe una mayor variación en la dieta. Pteranodontia contenía muchos taxones piscívoros, como Ornithocheirae , Boreopteridae , Pteranodontidae y Nyctosauridae. La partición del nicho hizo que los ornitoqueres y los nictosáuridos posteriores se alimentaran por inmersión aérea como las fragatas de hoy (con la excepción del Alcione elainus , adaptado para el buceo ), mientras que los boreopteridos eran animales buceadores de agua dulce similares a los cormoranes , y los pteranodontes buzos pelágicos similares a los piqueros. y alcatraces . Los istiodactylids probablemente eran principalmente carroñeros. [175] Archaeopterodactyloidea obtenía alimento en hábitats costeros o de agua dulce. Germanodactylus y Pterodactylus eran piscívoros, mientras que Ctenochasmatidae se alimentaban en suspensión, usando sus numerosos dientes finos para filtrar pequeños organismos de aguas poco profundas. Pterodaustro fue adaptado para la alimentación por filtración similar a un flamenco . [176]
En contraste, los Azhdarchoidea eran en su mayoría pterosaurios terrestres. Los tapejaridae eran omnívoros arbóreos que complementaban semillas y frutos con pequeños insectos y vertebrados. [166] [177] Dsungaripteridae eran moluscívoros especializados, que usaban sus poderosas mandíbulas para aplastar las conchas de moluscos y crustáceos. Los Thalassodromidae probablemente eran carnívoros terrestres. El propio Thalassodromeus recibió su nombre de un método de pesca conocido como "alimentación desnatada", que más tarde se entendió como biomecánicamente imposible. Quizás persiguió presas relativamente grandes, en vista de las articulaciones de sus mandíbulas reforzadas y la fuerza de mordida relativamente alta. [178] Azhdarchidaeahora se entiende que son depredadores terrestres similares a los cálaos terrestres o algunas cigüeñas , que comen cualquier presa que pudieran tragar entera. [179] Hatzegopteryx era un depredador de constitución robusta de presas relativamente grandes, incluidos dinosaurios de tamaño mediano. [180] [181] Alanqa pudo haber sido un especialista en moluscos. [182] Un asombroso estudio fue publicado en la revista Cretaceous Research. Los paleontólogos estaban analizando la mandíbula de una especie de pterosaurio Lonchodraco giganteus y encontraron foramen cerca de la punta de su pico y tenían un patrón similar al de las aves existentes, lo que sugeriría un hábito de alimentación táctil en los pterosaurios. [183]
Depredadores naturales
Se sabe que los terópodos se comieron a los pterosaurios . En la edición del 1 de julio de 2004 de Nature , el paleontólogo Eric Buffetaut analiza un fósil del Cretácico Inferior de tres vértebras cervicales de un pterosaurio con el diente roto de un espinosaurio , probablemente Irritator , incrustado en él. Se sabe que las vértebras no han sido comidas y expuestas a la digestión, ya que las articulaciones todavía están articuladas. [184]
Reproducción e historia de vida
Si bien se sabe muy poco sobre la reproducción de los pterosaurios, se cree que, al igual que todos los dinosaurios, todos los pterosaurios se reproducen poniendo huevos, aunque estos hallazgos son muy raros. El primer huevo de pterosaurio conocido se encontró en las canteras de Liaoning, el mismo lugar que produjo dinosaurios emplumados. El huevo estaba aplastado sin signos de agrietarse, por lo que evidentemente los huevos tenían cáscaras correosas, como en los lagartos modernos. [185] Esto fue respaldado por la descripción de un huevo de pterosaurio adicional perteneciente al género Darwinopterus , descrito en 2011, que también tenía una cáscara coriácea y, también como los reptiles modernos pero a diferencia de las aves, era bastante pequeño en comparación con el tamaño de la madre. . [186] En 2014, cinco huevos sin aplanar de la especie.Hamipterus tianshanensis se encontraron en un depósito del Cretácico Inferior en el noroeste de China. El examen de las cáscaras mediante microscopía electrónica de barrido mostró la presencia de una capa fina de cáscara de huevo calcárea con una membrana debajo. [187] Un estudio de la estructura y la química de la cáscara de huevo de los pterosaurios publicado en 2007 indicó que es probable que los pterosaurios hayan enterrado sus huevos, como los cocodrilos y las tortugas modernos. El entierro de huevos habría sido beneficioso para la evolución temprana de los pterosaurios, ya que permite más adaptaciones para reducir el peso, pero este método de reproducción también habría puesto límites a la variedad de entornos en los que los pterosaurios podrían vivir, y puede haberlos puesto en desventaja cuando empezaron a enfrentarse a la competencia ecológica de las aves. [188]
Un espécimen de Darwinopterus muestra que al menos algunos pterosaurios tenían un par de ovarios funcionales , a diferencia del ovario funcional único en las aves, descartando la reducción de los ovarios funcionales como un requisito para el vuelo motorizado. [189]
Las membranas de las alas conservadas en los embriones de pterosaurios están bien desarrolladas, lo que sugiere que los pterosaurios estaban listos para volar poco después del nacimiento. [190] Sin embargo, las tomografías de huevos de Hamipterus fosilizados sugieren que los pterosaurios jóvenes tenían huesos del muslo bien desarrollados para caminar, pero tórax débiles para volar. [191] Se desconoce si esto es cierto para otros pterosaurios. Se han encontrado fósiles de pterosaurios de sólo unos pocos días a una semana de edad (llamados "flapings"), que representan a varias familias de pterosaurios, incluidos los pterodáctílidos, los ramporinquidos, los ctenochasmátidos y los azdárquidos. [23] Todos conservan huesos que muestran un grado relativamente alto de endurecimiento ( osificación) para su edad y proporciones de alas similares a las de los adultos. De hecho, muchos pterosaurios aleteados se han considerado adultos y se han colocado en especies separadas en el pasado. Además, los flapings se encuentran normalmente en los mismos sedimentos que los adultos y los juveniles de la misma especie, como los flapings de Pterodactylus y Rhamphorhynchus que se encuentran en la caliza Solnhofen de Alemania y los flapings de Pterodaustro de Argentina. Todos se encuentran en ambientes acuáticos profundos lejos de la costa. [192]
Para la mayoría de las especies de pterosaurios, no se sabe si practicaron algún tipo de cuidado parental, pero su capacidad para volar tan pronto como emergieron del huevo y las numerosas aletas encontradas en ambientes alejados de los nidos y junto a los adultos ha llevado a la mayoría de los investigadores. , incluidos Christopher Bennett y David Unwin, para concluir que las crías dependieron de sus padres durante un período de tiempo relativamente corto, durante un período de rápido crecimiento, mientras que las alas crecieron lo suficiente para volar, y luego dejaron el nido para valerse por sí mismos. , posiblemente a los pocos días de la eclosión. [23] [193] Alternativamente, pueden haber usado productos de yema almacenada para alimentarse durante sus primeros días de vida, como en los reptiles modernos, en lugar de depender de los padres para alimentarse. [192] FosilizadoSe mostró que los nidos de Hamipterus conservaban muchos pterosaurios machos y hembras junto con sus huevos de una manera similar a la de las colonias modernas de aves marinas . [187] [194] Debido a lo poco desarrollado que estaban los cofres de las crías para volar, se sugirió que Hamipterus pudo haber practicado alguna forma de cuidado parental. [191] Sin embargo, este estudio ha sido criticado desde entonces. [195] La mayoría de la evidencia actualmente se inclina hacia que las crías de pterosaurio sean superprecociales , similar a la del megapodo.aves, que vuelan después de la eclosión sin necesidad del cuidado de sus padres. Un estudio adicional compara la evidencia de superprecocialidad y "vuelo tardío" y sugiere abrumadoramente que la mayoría, si no todos, los pterosaurios fueron capaces de volar poco después de la eclosión. [196]
Las tasas de crecimiento de los pterosaurios una vez que nacieron variaron entre los diferentes grupos. En los pterosaurios de cola larga más primitivos (" rhamphorhynchoids "), como Rhamphorhynchus , la tasa de crecimiento promedio durante el primer año de vida fue de 130% a 173%, un poco más rápido que la tasa de crecimiento de los caimanes . El crecimiento de estas especies se desaceleró después de la madurez sexual, y Rhamphorhynchus habría tardado más de tres años en alcanzar el tamaño máximo. [193] En contraste, los grandes pterosaurios pterodactiloides más avanzados, como el Pteranodon , crecieron hasta alcanzar el tamaño adulto durante el primer año de vida. Además, los pterodactiloides tenían un crecimiento determinado, lo que significa que los animales alcanzaron un tamaño adulto máximo fijo y dejaron de crecer. [192]
Patrones de actividad diaria
Se han utilizado comparaciones entre los anillos esclerales de los pterosaurios y las aves y reptiles modernos para inferir los patrones de actividad diaria de los pterosaurios. Se ha inferido que los géneros de pterosaurios Pterodactylus , Scaphognathus y Tupuxuara son diurnos , se ha inferido que Ctenochasma , Pterodaustro y Rhamphorhynchus son nocturnos , y Tapejara se ha inferido que es catemeral , estando activo durante el día durante cortos intervalos. Como resultado, Ctenochasma y Rhamphorhynchus , que posiblemente se alimentan de pecespuede haber tenido patrones de actividad similares a los de las aves marinas nocturnas modernas, y el Pterodaustro, que se alimenta por filtración, puede haber tenido patrones de actividad similares a los de las aves anseriformes modernas que se alimentan de noche. Las diferencias entre los patrones de actividad de los pterosaurios Ctenochasma , Rhamphorhynchus , Scaphognathus y Pterodactylus de Solnhofen también pueden indicar una división de nichos entre estos géneros. [197]
Importancia cultural
Los pterosaurios han sido un elemento básico de la cultura popular durante tanto tiempo como sus primos los dinosaurios, aunque por lo general no aparecen de manera prominente en películas, literatura u otro arte. Si bien la descripción de los dinosaurios en los medios populares ha cambiado radicalmente en respuesta a los avances en la paleontología, una imagen principalmente obsoleta de los pterosaurios ha persistido desde mediados del siglo XX. [198]
El término genérico vago "pterodáctilo" se usa a menudo para estas criaturas. Los animales representados con frecuencia representan Pteranodon o Rhamphorhynchus (no pterodactiloide) , o un híbrido ficticio de los dos. [198] Muchos juguetes y dibujos animados para niños presentan "pterodáctilos" con crestas parecidas a Pteranodon y largas colas y dientes parecidos a Rhamphorhynchus , una combinación que nunca existió en la naturaleza. Sin embargo, al menos un pterosaurio hizo tener tanto el Pteranodon cresta y los dientes -como: ludodactylus , cuyo nombre significa "juguete del dedo" por su parecido con los juguetes viejos, los niños inexactas. [199]Los pterosaurios a veces se han identificado incorrectamente como (los antepasados de) aves , aunque las aves son dinosaurios terópodos y no descendientes de pterosaurios.
Los pterosaurios se utilizaron en la ficción en la novela de 1912 de Sir Arthur Conan Doyle El mundo perdido y su adaptación cinematográfica de 1925 . Aparecieron en una serie de películas y programas de televisión desde entonces, incluida la película de 1933 King Kong y One Million Years BC de 1966.En este último, el animador Ray Harryhausen tuvo que agregar dedos de alas de murciélago inexactos a sus modelos de stop motion para mantener las membranas no se desmoronan, aunque este error particular era común en el arte incluso antes de que se hiciera la película. Rodan , un monstruo gigante ficticio (o kaiju ) que apareció por primera vez en la película Rodan de 1956 , es retratado como una enorme especie irradiada dePteranodon . [200] [201] Rodan ha aparecido en varias películas japonesas de Godzilla estrenadas durante las décadas de 1960, 1970, 1990 y 2000, y también apareció en la película de producción estadounidense de 2019 Godzilla: King of the Monsters . [202] [201]
Después de la década de 1960, los pterosaurios permanecieron en su mayoría ausentes de las apariciones notables en películas estadounidenses hasta Jurassic Park III de 2001 . El paleontólogo Dave Hone señaló que los pterosaurios en esta película no se habían actualizado significativamente para reflejar la investigación moderna. Los errores que persistieron fueron los dientes, mientras que el Pteranodon desdentado estaba destinado a ser representado, el comportamiento de anidación que se sabía que era inexacto en 2001 y las alas correosas, en lugar de las membranas tensas de la fibra muscular necesarias para el vuelo del pterosaurio. [198]
En la mayoría de las apariciones en los medios, los pterosaurios se representan como piscívoros , sin reflejar su variación dietética completa. También se muestran a menudo como depredadores aéreos similares a las aves de presa , agarrando víctimas humanas con garras en sus pies. Sin embargo, sólo se sabe que el pequeño anurognátido Vesperopterylus y el pequeño wukongopterido Kunpengopterus [203] poseen pies y manos prensiles, respectivamente; todos los demás pterosaurios tienen pies planos plantígrados sin dedos opuestos, y los pies son generalmente proporcionalmente pequeños, al menos en el caso de la Pteranodoncia. [204]
Ver también
- Animales voladores y planeadores
- Cronología gráfica de pterosaurios
- Lista de géneros de pterosaurios
- Filogenia de los pterosaurios
- Playa de pterosaurio
- Tamaño de pterosaurio
- Cronología de la investigación de pterosaurios
Notas
- ^ Adaptado de Witton (2013). [3] Grupos taxonómicos basados en Unwin et al. (2010). [4]
- ^ Ver la expansión de nichos ecológicos en el Mesozoico
Referencias
- ^ a b Andrés, B .; Clark, J .; Xu, X. (2014). "El pterodactiloide más antiguo y el origen del grupo" . Biología actual . 24 (9): 1011–16. doi : 10.1016 / j.cub.2014.03.030 . PMID 24768054 .
- ^ Barón, Matthew G. (2020). "Prueba de las relaciones entre grupos de pterosaurios a través de un muestreo más amplio de taxones y caracteres avemetatarsianos y una variedad de técnicas de análisis filogenético" . PeerJ . 8 : e9604. doi : 10.7717 / peerj.9604 . PMC 7512134 . PMID 33005485 .
- ^ Mark P. Witton (2013), Pterosaurios: Historia natural, Evolución, Anatomía , Princeton University Press, ISBN 978-0-691-15061-1
- ^ David M. Unwin (2010), " Darwinopterus y sus implicaciones para la filogenia del pterosaurio", Acta Geoscientica Sinica , 31 (1): 68-69
- ^ Jones, Daniel (2003) [1917], Peter Roach; James Hartmann; Jane Setter (eds.), Diccionario de pronunciación en inglés , Cambridge: Cambridge University Press, ISBN 978-3-12-539683-8
- ^ "Pterosaurio" . Diccionario Merriam-Webster .
- ^ "Distribución de pterosaurios en el tiempo y el espacio: un atlas" (PDF) . Zitteliana : 61–107. 2008.
- ^ Elgin RA, Hone DW, Frey E (2011). "La extensión de la membrana de vuelo del pterosaurio" . Acta Palaeontologica Polonica . 56 (1): 99-111. doi : 10.4202 / app.2009.0145 .
- ^ "Pterosaur.net :: Locomoción terrestre" . pterosaur.net . Consultado el 1 de febrero de 2020 .
- ^ Geggel 2018-12-17T19: 23: 17Z, Laura. "Es oficial: esos reptiles voladores llamados pterosaurios estaban cubiertos de plumas mullidas" . livescience.com . Consultado el 1 de febrero de 2020 .
- ^ a b Wang, X .; Kellner, AWA; Zhou, Z .; Campos, DA (2008). "Descubrimiento de un raro reptil volador arbóreo que habita en los bosques (Pterosauria, Pterodactyloidea) de China" . Actas de la Academia Nacional de Ciencias . 105 (6): 1983–87. Código Bibliográfico : 2008PNAS..105.1983W . doi : 10.1073 / pnas.0707728105 . PMC 2538868 . PMID 18268340 .
- ^ Lawson DA (marzo de 1975). "Pterosaurio del último Cretácico del oeste de Texas: descubrimiento de la criatura voladora más grande". Ciencia . 187 (4180): 947–948. Código Bibliográfico : 1975Sci ... 187..947L . doi : 10.1126 / science.187.4180.947 . PMID 17745279 . S2CID 46396417 .
- ^ Buffetaut E, Grigorescu D, Csiki Z (abril de 2002). "Un nuevo pterosaurio gigante con un cráneo robusto del último cretáceo de Rumania". Naturwissenschaften . 89 (4): 180–84. Código Bibliográfico : 2002NW ..... 89..180B . doi : 10.1007 / s00114-002-0307-1 . PMID 12061403 . S2CID 15423666 .
- ^ Benton, Michael J. (2004). "Origen y relaciones de Dinosauria". En Weishampel, David B .; Dodson, Peter; Osmólska, Halszka (eds.). La Dinosauria (2ª ed.). Berkeley: Prensa de la Universidad de California. pp. 7 -19. ISBN 978-0-520-24209-8.
- ^ a b Naish, Darren. "Pterosaurios: mitos y conceptos erróneos" . Pterosaur.net . Consultado el 18 de junio de 2011 .
- ↑ a b Alexander, David E. y Vogel, Steven (2004). Flyers de la naturaleza: pájaros, insectos y la biomecánica del vuelo . Prensa JHU. pag. 191. ISBN 978-0-8018-8059-9.
- ↑ a b c d Witmer LM, Chatterjee S, Franzosa J, Rowe T (2003). "Neuroanatomía de reptiles voladores e implicaciones para el vuelo, la postura y el comportamiento". Naturaleza . 425 (6961): 950–53. Código Bibliográfico : 2003Natur.425..950W . doi : 10.1038 / nature02048 . PMID 14586467 . S2CID 4431861 .
- ^ "Pterosaur.net :: Orígenes y relaciones" . pterosaur.net . Consultado el 1 de febrero de 2020 .
- ^ Andrés, Brian; Clark, James M .; Xing, Xu (29 de enero de 2010). "Un nuevo pterosaurio ramphorhynchid del Jurásico Superior de Xinjiang, China, y las relaciones filogenéticas de los pterosaurios basales" (PDF) . Revista de Paleontología de Vertebrados . 30 (1): 163–87. doi : 10.1080 / 02724630903409220 . ISSN 0272-4634 . S2CID 53688256 .
- ^ Witton, Mark P .; Martill, David M .; Loveridge, Robert F. (2010). "Cortar las alas de pterosaurios gigantes: comentarios sobre la diversidad y las estimaciones de envergadura". Acta Geoscientica Sinica . 31 : 79–81.
- ↑ Witton , 2013 , p. 58.
- ↑ a b c Witton , 2013 , p. 23.
- ↑ a b c d e f g Unwin, David M. (2006). Los pterosaurios: desde el tiempo profundo . Nueva York: Pi Press. pag. 246. ISBN 978-0-13-146308-0.
- ↑ a b Witton , 2013 , p. 27.
- ↑ a b Wellnhofer , 1991 , p. 47.
- ↑ a b c Witton , 2013 , p. 26.
- ↑ a b c Witton , 2013 , p. 24.
- ↑ a b Frey E, Martill DM (1998). "Conservación de tejidos blandos en un espécimen de Pterodactylus kochi (Wagner) del Jurásico Superior de Alemania". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen . 210 (3): 421–41. doi : 10.1127 / njgpa / 210/1998/421 .
- ^ Wellnhofer 1991 , p. 48.
- ↑ a b c d Naish D, Martill DM (2003). "Pterosaurios - una exitosa invasión de los cielos prehistóricos". Biólogo . 50 (5): 213–16.
- ↑ a b Czerkas, SA y Ji, Q. (2002). Un nuevo ramphorhynchoid con un cabecero y estructuras tegumentarias complejas. En: Czerkas, SJ (Ed.). Dinosaurios emplumados y el origen del vuelo . The Dinosaur Museum: Blanding, Utah, 15–41. ISBN 1-932075-01-1 .
- ^ Wellnhofer 1991 , p. 49.
- ^ S. Christopher Bennett (1994). "Taxonomía y sistemática del pterosaurio del Cretácico tardío Pteranodon (Pterosauria, Pterodactyloidea)" . Documentos ocasionales del Museo de Historia Natural de la Universidad de Kansas . 169 : 1-70.
- ↑ a b Witton , 2013 , p. 28.
- ↑ a b c Wellnhofer , 1991 , p. 50.
- ↑ Witton , 2013 , p. 45.
- ↑ a b c d Witton , 2013 , p. 46.
- ↑ a b c Witton , 2013 , p. 30.
- ↑ a b c Witton , 2013 , p. 31.
- ↑ a b Wellnhofer , 1991 , p. 51.
- ↑ a b c d e Wellnhofer , 1991 , p. 52.
- ↑ Witton , 2013 , p. 44.
- ↑ a b c Witton , 2013 , p. 32.
- ↑ a b Witton , 2013 , p. 54.
- ↑ a b Witton , 2013 , p. 53.
- ^ Bennett SC (2000). "Vuelo de pterosaurio: el papel de las actinofibrillas en la función de las alas". Biología histórica . 14 (4): 255–84. doi : 10.1080 / 10292380009380572 . S2CID 85185457 .
- ^ a b c Kellner, AWA; Wang, X .; Tischlinger, H .; Campos, D .; Hone, DWE; Meng, X. (2009). "El tejido blando de Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) y la estructura de la membrana del ala del pterosaurio" . Proceedings of the Royal Society B . 277 (1679): 321–29. doi : 10.1098 / rspb.2009.0846 . PMC 2842671 . PMID 19656798 .
- ↑ a b Claessens LP, O'Connor PM, Unwin DM (2009). Sereno P (ed.). "La evolución respiratoria facilitó el origen del vuelo de los pterosaurios y el gigantismo aéreo" . PLOS ONE . 4 (2): e4497. Código Bibliográfico : 2009PLoSO ... 4.4497C . doi : 10.1371 / journal.pone.0004497 . PMC 2637988 . PMID 19223979 .
- ↑ a b Witton , 2013 , p. 52.
- ↑ a b Witton , 2013 , p. 55.
- ↑ a b Unwin DM, Bakhurina NN (1994). " Sordes pilosus y la naturaleza del aparato de vuelo del pterosaurio". Naturaleza . 371 (6492): 62–64. Código Bibliográfico : 1994Natur.371 ... 62U . doi : 10.1038 / 371062a0 . S2CID 4314989 .
- ↑ Wang X, Zhou Z, Zhang F, Xu X (2002). "Un pterosaurio ramphorynchoid casi completamente articulado con membranas de alas y" pelos "excepcionalmente bien conservados de Mongolia Interior, noreste de China". Boletín de ciencia china . 47 (3): 3. Código Bibliográfico : 2002ChSBu..47..226W . doi : 10.1360 / 02tb9054 . S2CID 86641794 .
- ^ Frey, E .; Tischlinger, H .; Buchy, M.-C .; Martill, DM (2003). "Nuevos ejemplares de Pterosauria (Reptilia) con partes blandas con implicaciones para la anatomía y la locomoción de los pterosaurios". Sociedad Geológica, Londres, Publicaciones especiales . 217 (1): 233–66. Código Bibliográfico : 2003GSLSP.217..233F . doi : 10.1144 / GSL.SP.2003.217.01.14 . S2CID 130462931 .
- ^ Dyke GJ, Nudds RL, Rayner JM (julio de 2006). "Disparidad de extremidades y forma de alas en pterosaurios" . J. Evol. Biol . 19 (4): 1339–42. doi : 10.1111 / j.1420-9101.2006.01096.x . PMID 16780534 . S2CID 30516133 .
- ↑ a b c Wellnhofer , 1991 , p. 53.
- ↑ Witton , 2013 , p. 33.
- ↑ Witton , 2013 , p. 34.
- ^ Wilkinson MT, Unwin DM, Ellington CP (2006). "Función de elevación alta del hueso pteroide y el ala delantera de los pterosaurios" . Proceedings of the Royal Society B . 273 (1582): 119–26. doi : 10.1098 / rspb.2005.3278 . PMC 1560000 . PMID 16519243 .
- ^ Bennett SC (2007). "Articulación y función del hueso de pteroides de pterosaurios". Revista de Paleontología de Vertebrados . 27 (4): 881–91. doi : 10.1671 / 0272-4634 (2007) 27 [881: AAFOTP] 2.0.CO; 2 .
- ^ Zhou, Chang-Fu; Schoch, Rainer R. (2011). "Nuevo material del pterosaurio no pterodactiloide Changchengopterus pani Lü, 2009 de la Formación Tiaojishan del Jurásico Tardío de Liaoning occidental". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen . 260 (3): 265–75. doi : 10.1127 / 0077-7749 / 2011/0131 .
- ^ Wang, Xiao-Lin; Kellner, Alexander WA; Jiang, Shun-Xing; Cheng, Xin; Meng, Xi; Rodrigues, Taissa (2010). "Nuevos pterosaurios de cola larga (Wukongopteridae) del oeste de Liaoning, China" . Anais da Academia Brasileira de Ciências . 82 (4): 1045–62. doi : 10.1590 / s0001-37652010000400024 . PMID 21152776 .
- ^ Wilkinson MT; Unwin DM; Ellington CP (2006). "Función de elevación alta del hueso pteroide y el ala delantera de los pterosaurios" . Proceedings of the Royal Society B . 273 (1582): 119–26. doi : 10.1098 / rspb.2005.3278 . PMC 1560000 . PMID 16519243 .
- ↑ a b c d e f g h Witton , 2013 , pág. 35.
- ↑ a b c d Wellnhofer , 1991 , p. 55.
- ^ Wellnhofer 1991 , págs. 53–54.
- ↑ a b c d e f Wellnhofer 1991 , p. 56.
- ↑ a b c d e f Wellnhofer 1991 , p. 57.
- ↑ a b c Witton , 2013 , p. 36.
- ↑ a b c d Witton MP, Naish D (2008). McClain CR (ed.). "Una reevaluación de la morfología funcional del pterosaurio azdárquido y la paleoecología" . PLOS ONE . 3 (5): e2271. Código Bibliográfico : 2008PLoSO ... 3.2271W . doi : 10.1371 / journal.pone.0002271 . PMC 2386974 . PMID 18509539 .
- ↑ Witton , 2013 , p. 37.
- ↑ Witton , 2013 , p. 39.
- ↑ Witton , 2013 , p. 43.
- ↑ Witton , 2013 , p. 47.
- ↑ Witton , 2013 , p. 48.
- ↑ a b c d e Witton , 2013 , p. 51.
- ^ Goldfuss, A (1831). "Beiträge zur Erkentniss verschiedner Reptilien der Vorwelt". Nova Acta Academiae Leopoldinae . 15 : 61-128.
- ^ Benton, Michael J .; Xu, Xing; Orr, Patrick J .; Kaye, Thomas G .; Pittman, Michael; Kearns, Stuart L .; McNamara, Maria E .; Jiang, Baoyu; Yang, Zixiao (enero de 2019). "Estructuras tegumentarias de pterosaurio con ramificaciones complejas en forma de plumas" . Ecología y evolución de la naturaleza . 3 (1): 24–30. doi : 10.1038 / s41559-018-0728-7 . hdl : 1983 / 1f7893a1-924d-4cb3-a4bf-c4b1592356e9 . ISSN 2397-334X . PMID 30568282 . S2CID 56480710 .
- ↑ Briggs, Helen (17 de diciembre de 2018). "La piel vuela sobre nuevos fósiles de pterosaurios" . BBC News . Consultado el 19 de diciembre de 2018 .
- ^ Unwin & Martill (28 de septiembre de 2020). "No hay protoplumas en pterosaurios" . Ecología y Evolución de la Naturaleza . 4 (12): 1590-1591. doi : 10.1038 / s41559-020-01308-9 . PMID 32989266 .
- ^ Kellner; et al. (2009). "El tejido blando de Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) y la estructura de la membrana del ala del pterosaurio" . ResearchGate . 277 (1679): 321–29. doi : 10.1098 / rspb.2009.0846 . PMC 2842671 . PMID 19656798 .
- ^ Unwin, David M .; Bakhurina, Natasha N. (septiembre de 1994). "Sordes pilosus y la naturaleza del aparato de vuelo del pterosaurio" . Naturaleza . 371 (6492): 62–64. Código Bib : 1994Natur.371 ... 62U . doi : 10.1038 / 371062a0 . ISSN 1476-4687 . S2CID 4314989 .
- ^ Yang, Zixiao; Jiang, Baoyu; McNamara, Maria E .; Kearns, Stuart L .; Pittman, Michael; Kaye, Thomas G .; Orr, Patrick J .; Xu, Xing; Benton, Michael J. (28 de septiembre de 2020). "Respuesta a: No hay protoplumas en pterosaurios" . Ecología y evolución de la naturaleza . 4 (12): 1592-1593. doi : 10.1038 / s41559-020-01309-8 . ISSN 2397-334X . PMID 32989267 .
- ↑ a b Witton , 2013 , p. 5.
- ^ Wellnhofer 1991 , p. 22.
- ↑ Witton , 2013 , p. 6.
- ^ Witton 2013 , págs. 6-7.
- ↑ a b c d Witton , 2013 , p. 7.
- ↑ Collini, CA (1784). "Sur quelques Zoolithes du Cabinet d'Histoire naturelle de SASE Palatine & de Bavière, à Mannheim". Acta Theodoro-Palatinae Mannheim 5 Pars Physica , págs. 58-103 (1 lámina).
- ^ Wagler, J. (1830). Natürliches System der Amphibien Munich, 1830: 1-354.
- ↑ Cuvier G (1801). "[Reptile volant]. En: Extrait d'un ouvrage sur les espèces de quadrupèdes dont on a trouvé les ossemens dans l'intérieur de la terre". Journal de Physique, de Chimie et d'Histoire Naturelle . 52 : 253–67.
- ^ Cuvier, G., 1809, "Mémoire sur le squelette fossile d'un Reptil volant des environs d'Aichstedt, que quelques naturalistes ont pris pour un oiseau, et donc nous formons un genre de Sauriens, sous le nom de Ptero-Dactyle ", Annales du Musée d'Histoire Naturelle , París, 13 págs. 424–37
- ^ Rafinesque, CS, 1815, Analyse de la Nature ou tableau de l'univers et des corps organisés , Palermo
- ↑ Von Soemmerring, ST, 1812, "Über einen Ornithocephalus oder über das unbekannten Thier der Vorwelt, dessen Fossiles Gerippe Collini im 5. in der Naturalien-Sammlung der königlichen Akademie der Wissenschaften zu München befindet ", Denkschriften der königlichen bayerischen Akademie der Wissenschaften , München: math-physikalische Classe 3 : 89–158
- ^ Wellnhofer 1991 , p. 27.
- ^ Newman, E (1843). "Nota sobre la tribu Pterodactyle considerada como murciélagos marsupiales". Zoólogo . 1 : 129–31.
- ↑ Kaup, J (1834). "Versuch einer Eintheilung der Säugethiere en 6 Stämme und der Amphibien en 6 Ordnungen". Isis von Oken . 1834 : 315.
- ^ Wellnhofer 1991 , p. 28.
- ^ Wellnhofer 1991 , p. 29.
- ^ Wellnhofer 1991 , p. 33.
- ^ Seeley, HG, 1870, Ornithosauria - un estudio elemental de los huesos de Pterodactyles , Cambridge University Press
- ↑ Seeley, HG, 1901, Dragons of the Air: An cuenta de reptiles voladores extintos , Londen: Methuen
- ↑ Mivart, G (1881). "Un relato popular de camaleones". Naturaleza . 24 (615): 309–38. Código bibliográfico : 1881Natur..24..335. . doi : 10.1038 / 024335f0 . S2CID 30819954 .
- ↑ a b Wellnhofer , 1991 , p. 35.
- ↑ a b c Wellnhofer , 1991 , p. 36.
- ↑ a b Wellnhofer , 1991 , p. 31.
- ^ Wellnhofer 1991 , págs. 37–38.
- ^ Marsh, OC (1882). "Las alas de los pterodáctilos". Revista estadounidense de ciencia . 3 (16): 223.
- ↑ Zittel, KA (1882). "Über Flugsaurier aus dem lithografischen Schiefer Bayerns". Palaeontographica . 29 : 47–80.
- ^ Broili, F., 1927, "Ein Ramphorhynchus mit Spuren von Haarbedeckung", Sitzungsberichte der Bayerischen Akademie der Wissenschaften p. 49-67
- ↑ Edinger, T (1927). "Das Gehirn der Pterosaurier" (PDF) . Zeitschrift für Anatomie und Entwicklungsgeschichte . 83 (1/3): 105-12. doi : 10.1007 / bf02117933 . S2CID 19084773 . Archivado desde el original (PDF) el 28 de julio de 2020 . Consultado el 27 de octubre de 2019 .
- ^ Hankin EH y Watson DSM; "Sobre el vuelo de los pterodáctilos", The Aeronautical Journal , octubre de 1914, págs. 324–35
- ^ Bakker, Robert, 1986, Las herejías de los dinosaurios , Londen: Penguin Books, 1988, p. 283
- ^ Padian, K (1979). "Las alas de los pterosaurios: una nueva mirada". Descubrimiento . 14 : 20-29.
- ^ Padian, K., 1980, Estudios de la estructura, evolución y vuelo de los pterosaurios (reptilia: Pterosauria) , Ph.D. diss., Departamento de Biología, Universidad de Yale
- ↑ a b Padian K (1983). "Un análisis funcional de volar y caminar en pterosaurios". Paleobiología . 9 (3): 218–39. doi : 10.1017 / S009483730000765X . JSTOR 2400656 .
- ↑ a b c d e Witton , 2013 , p. 9.
- ^ Wellnhofer, P., 1978, Handbuch der Paläoherpetologie XIX. Pterosauria , Urban & Fischer, München
- ^ Wellnhofer 1991 , págs. 1-192.
- ^ Dyke, GJ McGowan; Nudds, RL; Smith, D. (2009). "La forma de la evolución de pterosaurios: evidencia del registro fósil" . Revista de Biología Evolutiva . 22 (4): 890–98. doi : 10.1111 / j.1420-9101.2008.01682.x . PMID 19210587 . S2CID 32518380 .
- ↑ a b Witton, 2013
- ↑ a b Witton , 2013 , p. 10.
- ↑ Witton , 2013 , p. 13.
- ^ Witton 2013 , págs.14, 17.
- ^ Benton, MJ (1999). " Scleromochlus taylori y el origen de dinosaurios y pterosaurios" . Transacciones filosóficas de la Royal Society B: Ciencias biológicas . 354 (1388): 1423–46. doi : 10.1098 / rstb.1999.0489 . PMC 1692658 .
- ^ Bennett, S. Christopher (1996). "La posición filogenética de la Pterosauria dentro de la Archosauromorpha" . Revista Zoológica de la Sociedad Linneana . 118 (3): 261-308. doi : 10.1111 / j.1096-3642.1996.tb01267.x .
- ^ Irmis, RB; Nesbitt, SJ; Padian, K .; Smith, ND; Turner, AH; Woody, D .; Downs, A. (2007). "Un ensamblaje de Dinosauromorfo del Triásico Tardío de Nuevo México y el surgimiento de los dinosaurios". Ciencia . 317 (5836): 358–61. Código bibliográfico : 2007Sci ... 317..358I . doi : 10.1126 / science.1143325 . PMID 17641198 . S2CID 6050601 .
- ^ a b Hone DWE; Benton MJ (2007). "Una evaluación de las relaciones filogenéticas de los pterosaurios a los reptiles arcosauromorfos". Revista de Paleontología Sistemática . 5 (4): 465–69. doi : 10.1017 / S1477201907002064 . S2CID 86145645 .
- ^ Nesbitt, SJ (2011). "La evolución temprana de los arcosaurios: relaciones y el origen de los principales clados" (PDF) . Boletín del Museo Americano de Historia Natural . 352 : 1–292. doi : 10.1206 / 352.1 . hdl : 2246/6112 . S2CID 83493714 .
- ↑ Ezcurra, Martín D. (28 de abril de 2016). "Las relaciones filogenéticas de los arcosauromorfos basales, con énfasis en la sistemática de los arcosauriformes proterosuquios" . PeerJ . 4 : e1778. doi : 10.7717 / peerj.1778 . ISSN 2167-8359 . PMC 4860341 . PMID 27162705 .
- ↑ Bennett, SC (2020). "Reevaluación del archosauriforme Triásico Scleromochlus taylori : ni corredor ni bípedo, sino saltador" . PeerJ . 8 : e8418. doi : 10.7717 / peerj.8418 . PMC 7035874 . PMID 32117608 .
- ↑ Ezcurra, Martín D .; Nesbitt, Sterling J .; Bronzati, Mario; Dalla Vecchia, Fabio Marco; Agnolin, Federico L .; Benson, Roger BJ; Brissón Egli, Federico; Cabreira, Sergio F .; Evers, Serjoscha W .; Gentil, Adriel R .; Irmis, Randall B. (9 de diciembre de 2020). "Los precursores de dinosaurios enigmáticos cierran la brecha con el origen de Pterosauria" . Naturaleza . 588 (7838): 445–449. Código Bib : 2020Natur.588..445E . doi : 10.1038 / s41586-020-3011-4 . ISSN 1476-4687 . PMID 33299179 . S2CID 228077525 .
- ^ "Los paleontólogos encuentran precursores de pterosaurios que llenan un vacío en la historia evolutiva temprana" . phys.org . Consultado el 14 de diciembre de 2020 .
- ^ Negro, Riley. "Pterosaur Origins Flap into Focus" . Scientific American . Consultado el 14 de diciembre de 2020 .
- ↑ Witton , 2013 , p. 18.
- ^ Rupert Wild, 1983, "Über die Ursprung der Flugsaurier", Weltenberger Akademie, Erwin Rutte-Festschrift , págs. 231–38
- ↑ a b c d Witton, Mark P. (2015). "¿Eran los primeros pterosaurios locomotoras terrestres ineptas?" . PeerJ . 3 : e1018. doi : 10.7717 / peerj.1018 . PMC 4476129 . PMID 26157605 .
- ^ Documental de la BBC: Caminando con dinosaurios (episodio 4) - Gigante de los cielos a los 22 ', Tim Haines, 1999
- ^ Slack KE, Jones CM, Ando T, et al. (Junio de 2006). "Los primeros fósiles de pingüinos, además de genomas mitocondriales, calibran la evolución aviar" . Biología Molecular y Evolución . 23 (6): 1144–55. doi : 10.1093 / molbev / msj124 . PMID 16533822 .
- ^ Mayordomo, Richard J .; Barrett, Paul M .; Nowbath, Stephen y Upchurch, Paul (2009). "Estimación de los efectos de los sesgos de muestreo en los patrones de diversidad de pterosaurios: implicaciones para las hipótesis de reemplazo competitivo de aves / pterosaurios". Paleobiología . 35 (3): 432–46. doi : 10.1666 / 0094-8373-35.3.432 . S2CID 84324007 .
- ^ a b Longrich, NR; Martill, DM; Andrés, B. (2018). "Pterosaurios tardíos de Maastrichtian del norte de África y extinción masiva de Pterosauria en el límite Cretácico-Paleógeno" . PLOS Biología . 16 (3): e2001663. doi : 10.1371 / journal.pbio.2001663 . PMC 5849296 . PMID 29534059 .
- ^ Andrés, B .; Myers, TS (2013). "Pterosaurios estrella solitaria". Transacciones de ciencias ambientales y de la tierra de la Royal Society of Edinburgh . 103 (3–4): 1. doi : 10.1017 / S1755691013000303 . S2CID 84617119 .
- ^ Agnolin, Federico L. y Varricchio, David (2012). "Reinterpretación sistemática de Piksi barbarulna Varricchio, 2002 de la formación Two Medicine (Cretácico superior) del oeste de EE. UU. (Montana) como un pterosaurio en lugar de un pájaro" (PDF) . Geodiversitas . 34 (4): 883–94. doi : 10.5252 / g2012n4a10 . S2CID 56002643 .
- ↑ Prondvai, E .; Bodor, ER; Ösi, A. (2014). "¿La morfología refleja la ontogenia basada en la osteohistología? Un estudio de caso de sinfisis de mandíbulas de pterosaurio del Cretácico tardío de Hungría revela una diversidad taxonómica oculta" (PDF) . Paleobiología . 40 (2): 288–321. doi : 10.1666 / 13030 . S2CID 85673254 .
- ^ Martin-Silverstone, Elizabeth; Witton, Mark P .; Arbor, Victoria M .; Currie, Philip J. (2016). "Un pequeño pterosaurio azdarcoide del último Cretácico, la era de los gigantes voladores" . Ciencia Abierta de la Royal Society . 3 (8): 160333. Código Bibliográfico : 2016RSOS .... 360333M . doi : 10.1098 / rsos.160333 . PMC 5108964 . PMID 27853614 .
- ↑ Haluza, A .; Apesteguía, S. (2007). "Restos de pterosaurios (Archosauria, Ornithodira) del Cretácico superior temprano de" La Buitrera ", Río Negro, Argentina" . XXIII Jornadas Argentinas de Paleontología de Vertebrados .
- ^ Kellner, AW (2003). "Filogenia de pterosaurios y comentarios sobre la historia evolutiva del grupo". Sociedad Geológica, Londres, Publicaciones especiales . 217 (1): 105–37. Código bibliográfico : 2003GSLSP.217..105K . doi : 10.1144 / gsl.sp.2003.217.01.10 . S2CID 128892642 .
- ^ Nesbitt, SJ, Desojo, JB e Irmis, RB (2013). Anatomía, filogenia y paleobiología de los primeros archosaurios y sus parientes . Sociedad Geológica de Londres. ISBN 1862393613
- ↑ de Queiroz, K .; Cantino, PD; Gauthier, JA, eds. (2020). Phylónimos: un compañero del PhyloCode . CRC Press Boca Raton, FL. pag. 2072. ISBN 9780429821202.
- ^ Padian, K. (1997). "Pterosauromorpha", págs. 617-18 en Currie, PJ y Padian, K. The Encyclopedia of Dinosaurs . Prensa académica. ISBN 0122268105 .
- ^ Lü J .; Unwin DM; Xu L .; Zhang X. (2008). "Un nuevo pterosaurio azdarcoide del Cretácico Inferior de China y sus implicaciones para la filogenia y la evolución del pterosaurio". Naturwissenschaften . 95 (9): 891–97. Código Bibliográfico : 2008NW ..... 95..891L . doi : 10.1007 / s00114-008-0397-5 . PMID 18509616 . S2CID 13458087 .
- ↑ a b Alleyne, Richard (1 de octubre de 2008). "Los pterodáctilos eran demasiado pesados para volar, afirma un científico" . El telégrafo . Archivado desde el original el 31 de octubre de 2009 . Consultado el 2 de marzo de 2012 .
- ^ Powell, Devin (2 de octubre de 2008). "¿Eran los pterosaurios demasiado grandes para volar?" . NewScientist . Consultado el 2 de marzo de 2012 .
- ^ Templin, RJ; Chatterjee, Sankar (2004). Postura, locomoción y paleoecología de pterosaurios . Boulder, Colo: Sociedad Geológica de América. pag. 60. ISBN 978-0-8137-2376-1.
- ^ Naish, Darren (18 de febrero de 2009). "Los pterosaurios respiraban como los pájaros y tenían sacos de aire inflables en sus alas" . ScienceBlogs . Archivado desde el original el 21 de febrero de 2009 . Consultado el 3 de abril de 2016 .
- ^ a b c "Por qué los pterosaurios no daban tanto miedo después de todo" . El periódico The Observer . 11 de agosto de 2013 . Consultado el 12 de agosto de 2013 .
- ↑ a b Hecht, Jeff (16 de noviembre de 2010). "¿Saltaron los pterosaurios gigantes como murciélagos vampiros?" . NewScientist . Consultado el 2 de marzo de 2012 .
- ^ MacCready, P. (1985). "El Gran Proyecto Pterodáctilo" (PDF) . Ingeniería y ciencia . 49 (2): 18-24.
- ^ Molotsky, Irvin (28 de enero de 1986). "Con alas batiendo, modelo Pterodactly toma el aire" . New York Times .
- ^ Geist, N .; Hillenius, W .; Frey, E .; Jones, T .; Elgin, R. (2014). "Respirar en una caja: restricciones en la ventilación pulmonar en pterosaurios gigantes" . El registro anatómico . 297 (12): 2233–53. doi : 10.1002 / ar.22839 . PMID 24357452 . S2CID 27659270 .
- ^ Hopson JA (1977). "Tamaño relativo del cerebro y comportamiento en reptiles archosaurianos". Revisión anual de ecología y sistemática . 8 : 429–48. doi : 10.1146 / annurev.es.08.110177.002241 .
- ^ Anthes, Emily (18 de noviembre de 2013). "Sangre fría no significa estúpido" . The New York Times .
- ^ Codorniú, Laura; Paulina Carabajal, Ariana; Pol, Diego; Unwin, David; Rauhut, Oliver WM (2016). "Un pterosaurio jurásico de la Patagonia y el origen del neurocráneo pterodactiloide" . PeerJ . 4 : e2311. doi : 10.7717 / peerj.2311 . PMC 5012331 . PMID 27635315 .
- ^ Padian K (2003). "Posición y marcha del pterosaurio y la interpretación de las vías". Ichnos . 10 (2–4): 115–26. doi : 10.1080 / 10420940390255501 . S2CID 129113446 .
- ^ Hwang K, Huh M, Lockley MG, Unwin DM, Wright JL (2002). "Nuevas huellas de pterosaurios (Pteraichnidae) de la formación Uhangri del Cretácico tardío, suroeste de Corea" . Revista geológica . 139 (4): 421–35. Código bibliográfico : 2002GeoM..139..421H . doi : 10.1017 / S0016756802006647 . S2CID 54996027 .
- ^ Unwin DM (1997). "Huellas de pterosaurios y la capacidad terrestre de los pterosaurios". Lethaia . 29 (4): 373–86. doi : 10.1111 / j.1502-3931.1996.tb01673.x .
- ↑ a b c Witton , 2013 , p. 51
- ^ Longrich, NR; Martill, DM; Andrés, B. (2018). "Pterosaurios tardíos de Maastrichtian del norte de África y extinción masiva de Pterosauria en el límite Cretácico-Paleógeno" . PLOS Biología . 16 (3): e2001663. doi : 10.1371 / journal.pbio.2001663 . PMC 5849296 . PMID 29534059 .
- ^ Bennett, SC (2007). "Un segundo espécimen del pterosaurio Anurognathus ammoni ". Paläontologische Zeitschrift . 81 (4): 376–98. doi : 10.1007 / bf02990250 . S2CID 130685990 .
- ↑ Witton , 2013 , p. 103.
- ↑ Witton , 2013 , p. 121.
- ↑ Witton , 2013 , p. 122.
- ^ Andrés, B .; Clark, JM; Xing, X. (2010). "Un nuevo pterosaurio ramphorhynchid del Jurásico Superior de Xinjiang, China, y las relaciones filogenéticas de los pterosaurios basales" (PDF) . Revista de Paleontología de Vertebrados . 30 (1): 163–87. doi : 10.1080 / 02724630903409220 . S2CID 53688256 .
- ↑ Witton , 2013 , p. 134.
- ^ Lü J .; Xu L .; Chang H .; Zhang X. (2011). "Un nuevo pterosaurio darwinopterid del Jurásico medio de Liaoning occidental, noreste de China y sus implicaciones ecológicas". Acta Geologica Sinica - Edición en inglés . 85 (3): 507-14. doi : 10.1111 / j.1755-6724.2011.00444.x .
- ^ Witton 2013 , págs. 150–51.
- ↑ Witton , 2013 , p. 199.
- ^ Wu, Wen-Hao; Zhou, Chang-Fu; Andrés, Brian (2017). "El pterosaurio desdentado Jidapterus edentus (Pterodactyloidea: Azhdarchoidea) del Cretácico temprano Jehol Biota y sus implicaciones paleoecológicas" . PLOS ONE . 12 (9): e0185486. Código bibliográfico : 2017PLoSO..1285486W . doi : 10.1371 / journal.pone.0185486 . PMC 5614613 . PMID 28950013 .
- ^ Pêgas, RV y Kellner, AW (2015). Reconstrucción miológica mandibular preliminar de Thalassodromeus sethi (Pterodactyloidea: Tapejaridae). Flugsaurier 2015 Portsmouth, resúmenes, 47–48
- ^ Witton, diputado; Naish, D. (2015). "Pterosaurios azdárquidos: imitadores de pelícanos rastreadores de agua o" acechadores terrestres "? . Acta Palaeontologica Polonica . 60 (3). doi : 10.4202 / app.00005.2013 .
- ^ Naish, D .; Witton, diputado (2017). "La biomecánica del cuello indica que los pterosaurios azdárquidos gigantes de Transilvania eran depredadores de arco de cuello corto" . PeerJ . 5 : e2908. doi : 10.7717 / peerj.2908 . PMC 5248582 . PMID 28133577 .
- ^ Witton, M .; Brusatte, S .; Dyke, G .; Naish, D .; Norell, M .; Vremir, M. (2013). Señores pterosaurios de Transilvania: azdárquidos gigantes de cuello corto en la Rumanía del Cretácico Superior . El Simposio Anual de Paleontología de Vertebrados y Anatomía Comparada. Edimburgo. Archivado desde el original el 6 de abril de 2016.
- ^ Martill, DM; Ibrahim, N. (2015). "Una modificación inusual de las mandíbulas en cf. Alanqa , un pterosaurio azdárquido del Cretácico medio de los lechos Kem Kem de Marruecos" (PDF) . Investigación del Cretácico . 53 : 59–67. doi : 10.1016 / j.cretres.2014.11.001 .
- ^ Martill, David M .; Smith, Roy E .; Longrich, Nicholas; Brown, James (3 de septiembre de 2020). "Evidencia de alimentación táctil en pterosaurios: una punta sensible al pico de Lonchodraco giganteus (Pterosauria, Lonchodectidae) del Cretácico superior del sur de Inglaterra" . Investigación del Cretácico . 117 : 104637. doi : 10.1016 / j.cretres.2020.104637 . ISSN 0195-6671 .
- ^ Buffetaut E, Martill D, Escuillié F (julio de 2004). "Pterosaurios como parte de una dieta de espinosaurios" . Naturaleza . 430 (6995): 33. Bibcode : 2004Natur.429 ... 33B . doi : 10.1038 / 430033a . PMID 15229562 . S2CID 4398855 .
- ^ Ji Q, Ji SA, Cheng YN, et al. (Diciembre de 2004). "Paleontología: huevo de pterosaurio con cáscara coriácea". Naturaleza . 432 (7017): 572. Código Bibliográfico : 2004Natur.432..572J . doi : 10.1038 / 432572a . PMID 15577900 . S2CID 4416203 .
- ^ Lü J .; Unwin DM; Considerándolo DC; Jin X .; Liu Y .; Ji Q. (2011). "Una asociación de huevo-adulto, género y reproducción en pterosaurios". Ciencia . 331 (6015): 321–24. Código Bibliográfico : 2011Sci ... 331..321L . doi : 10.1126 / science.1197323 . PMID 21252343 . S2CID 206529739 .
- ↑ a b Wang, Xiaolin (2014). "Pterosaurios sexualmente dimórficos preservados tridimensionalmente y sus huevos de China" . Biología actual . 24 (12): 1323–30. doi : 10.1016 / j.cub.2014.04.054 . PMID 24909325 . [ enlace muerto permanente ]
- ^ Grellet-Tinner G, Wroe S, Thompson MB, Ji Q (2007). "Una nota sobre el comportamiento de anidación de pterosaurios". Biología histórica . 19 (4): 273–77. doi : 10.1080 / 08912960701189800 . S2CID 85055204 .
- ^ Xiaolin Wang, Kellner Alexander WA; Cheng, Xin; Jiang, Shunxing; Wang, Qiang; Sayão Juliana, M .; Rordrigues Taissa, Costa Fabiana R .; Recubrimiento; Meng, Xi; Zhou, Zhonghe (2015). "La cáscara de huevo y la histología proporcionan información sobre la historia de vida de un pterosaurio con dos ovarios funcionales" . Anais da Academia Brasileira de Ciências . 87 (3): 1599–1609. doi : 10.1590 / 0001-3765201520150364 . PMID 26153915 .
- ^ Wang X, Zhou Z (junio de 2004). "Paleontología: embrión de pterosaurio del Cretácico temprano" . Naturaleza . 429 (6992): 621. Bibcode : 2004Natur.429..621W . doi : 10.1038 / 429621a . PMID 15190343 . S2CID 4428545 .
- ^ a b "Las crías de pterosaurio necesitaban a sus padres, revela un tesoro de huevos (Actualización)" . phys.org . Consultado el 21 de marzo de 2020 .
- ↑ a b c Bennett SC (1995). "Un estudio estadístico de Rhamphorhynchus de la piedra caliza de Solnhofen de Alemania: clases anuales de una sola especie grande". Revista de Paleontología . 69 (3): 569–80. doi : 10.1017 / S0022336000034946 . JSTOR 1306329 .
- ↑ a b Prondvai, E .; Stein, K .; Ősi, A .; Sander, diputado (2012). Soares, Daphne (ed.). "Historia de vida de Rhamphorhynchus inferida de la histología ósea y la diversidad de estrategias de crecimiento de pterosaurios" . PLOS ONE . 7 (2): e31392. Código bibliográfico : 2012PLoSO ... 731392P . doi : 10.1371 / journal.pone.0031392 . PMC 3280310 . PMID 22355361 .
- ^ "Primeros huevos de pterosaurio 3D encontrados con sus padres" . phys.org . Consultado el 21 de marzo de 2020 .
- ^ Unwin, David Michael; Deeming, D. Charles (2019). "Desarrollo prenatal en pterosaurios y sus implicaciones para su capacidad locomotora postnatal" . Actas de la Royal Society B: Ciencias Biológicas . 286 (1904). doi : 10.1098 / rspb.2019.0409 . PMC 6571455 . PMID 31185866 .
- ^ https://www.nature.com/articles/s41598-021-92499-z
- ↑ Schmitz, L .; Motani, R. (2011). "Nocturnidad en dinosaurios inferidos de la morfología de la órbita y el anillo escleral". Ciencia . 332 (6030): 705–08. Código bibliográfico : 2011Sci ... 332..705S . doi : 10.1126 / science.1200043 . PMID 21493820 . S2CID 33253407 .
- ↑ a b c Hone, D. (2010). "Pterosaurios en la cultura popular". Pterosaur.net , consultado el 27 de agosto de 2010.
- ^ Frey, E., Martill, D. y Buchy, M. (2003). "Un nuevo ornitoqueirido con cresta del Cretácico Inferior del noreste de Brasil y la muerte inusual de un pterosaurio inusual" en: Buffetaut, E. y Mazin, J.-M. (eds.). Evolución y paleobiología de los pterosaurios . Publicación especial de la Sociedad Geológica 217 : 56–63. ISBN 1-86239-143-2 .
- ^ Berry 2005 , p. 452.
- ↑ a b Thomas, HN (2020). "El nacido del fuego: un análisis pterosaurológico de Rodan" . Journal of Geek Studies 7 : 53–59.
- ^ Gonzales, Dave (12 de octubre de 2016). "Un desglose del tamaño de un monstruo de cada película loca de 'Godzilla'" . Emocionante . Consultado el 11 de julio de 2019 .; Sharf, Zack (10 de diciembre de 2018). " El tráiler de ' Godzilla: King of the Monsters' convierte a Mothra, Rodan y más en un espectáculo épico" (vídeo) . IndieWire . Consultado el 11 de julio de 2019 .
- ^ Zhou, X .; Pêgas, RV; Ma, W .; Colgar.; Jin, X .; Leal, MEC; Bonde, N .; Kobayashi, Y .; Lautenschlager, S .; Wei, X .; Shen, C .; Ji, S. (2021). "Un nuevo pterosaurio darwinopteran revela arboricultura y un pulgar opuesto" . Biología actual . 31 (11): 2429–2436.e7. doi : 10.1016 / j.cub.2021.03.030 . PMID 33848460 .
- ^ "Mitos y conceptos erróneos" . Pterosaur.net
Obras citadas
- Berry, Mark F. (2005). La filmografía de dinosaurios . McFarland & Company. ISBN 978-0786424535.
- Wellnhofer, Peter (1991). La enciclopedia ilustrada de pterosaurios: una historia natural ilustrada de los reptiles voladores de la era mesozoica . Libros Crescent. ISBN 0517037017.
- Witton, Mark (2013). Pterosaurios: historia natural, evolución, anatomía . Prensa de la Universidad de Princeton. ISBN 978-0691150611.
Enlaces externos
- Pterosaur.net , sitio web de varios autores sobre todos los aspectos de la ciencia de los pterosaurios
- La base de datos de pterosaurios , por Paul Pursglove
- "Comentarios sobre la filogenia de la pterodactyloidea" , por Alexander WA Kellner (técnico)
| vtmiAvemetatarsalia / Pan-Aves | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| vtmiPterosauromorpha | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



| vtmiAletas , extremidades y alas | ||
|---|---|---|
| Aletas |
|  |
| Extremidades |
| |
| Alas |
| |
| Evolución |
| |
| Relacionada |
|