| Centro de color | |
|---|---|
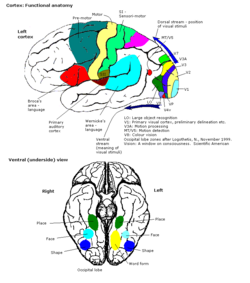 El área de visión de color se muestra como V8 en la imagen superior | |
| Terminología anatómica | |
El centro de color es una región del cerebro principalmente responsable de la percepción visual y el procesamiento cortical de las señales de color recibidas por el ojo, lo que finalmente da como resultado la visión de los colores . Se cree que el centro de color en los seres humanos está ubicado en el lóbulo occipital ventral como parte del sistema visual , además de otras áreas responsables de reconocer y procesar estímulos visuales específicos , como caras, palabras y objetos. Muchos estudios de imágenes de resonancia magnética funcional (fMRI) tanto en humanos como en monos macacos han demostrado que los estímulos de color activan múltiples áreas del cerebro, incluida la circunvolución fusiforme.y la circunvolución lingual . Estas áreas, así como otras identificadas por tener un papel en el procesamiento de la visión del color, se etiquetan colectivamente como área visual 4 ( V4 ). Los mecanismos exactos, la ubicación y la función de V4 aún se están investigando.
Corteza visual primaria
La parte principal de la corteza visual , (V1), se encuentra en el surco calcarino y es la primera área cortical involucrada en el procesamiento visual. Recibe información visual del núcleo geniculado lateral , que se encuentra en el tálamo . V1 envía la información visual recibida del LGN a otras áreas de la corteza extraestriada para un procesamiento de orden superior. Este procesamiento de orden superior incluye el reconocimiento de formas, movimiento y color. [1]
V1 tiene varias áreas que son sensibles al color, lo que indica que el procesamiento del color no se limita a un área. Según un artículo del Dr. Robert Shapley, V1 tiene un papel importante en la percepción del color. Los resultados experimentales de fMRI mostraron que V1 tiene dos tipos de neuronas sensibles al color: células de un solo oponente y células de doble oponente. Estas células son parte integral del proceso del oponente de interpretar las señales de color. Las neuronas de un solo oponente responden a grandes áreas de color. Esto es ventajoso para reconocer grandes ambientes y escenas de color. En comparación, las células oponentes dobles responden a patrones, texturas y límites de color. Esto es más importante para percibir el color de objetos e imágenes. Las células de doble oponente son receptivas a entradas opuestas de diferentes células de conoen la retina . Esto es ideal para identificar colores contrastantes, como rojo y verde. [1] Las células de doble oponente son particularmente importantes para calcular las proporciones de los conos locales a partir de la información visual de sus campos receptivos . [1] [2]
Las neuronas sensibles al color de un solo oponente se pueden dividir en dos categorías según las señales que reciben de las células del cono: neuronas LM y neuronas S / (L + M). Los tres tipos de células cónicas, pequeñas (S), medianas (M) y largas (L), detectan diferentes longitudes de onda en todo el espectro visible.. Las células del cono S pueden ver colores de longitud de onda corta, que corresponden al violeta y al azul. De manera similar, las celdas M detectan colores de longitud de onda media, como el verde y el amarillo, y las celdas L detectan colores de longitud de onda larga, como el rojo. Las neuronas LM, también llamadas células oponentes rojo-verde, reciben entrada de conos de longitud de onda larga opuestos por entrada de conos de longitud de onda media. Las neuronas S / (L + M) reciben información de las células S y se oponen a la suma de las entradas de las células L y M. Las neuronas S / (L + M) también se denominan células oponentes azul-amarillas. La oposición entre los colores permite que el sistema visual interprete las diferencias de color, lo que en última instancia es más eficiente que procesar los colores por separado. [1] [3]
Procesamiento visual de orden superior
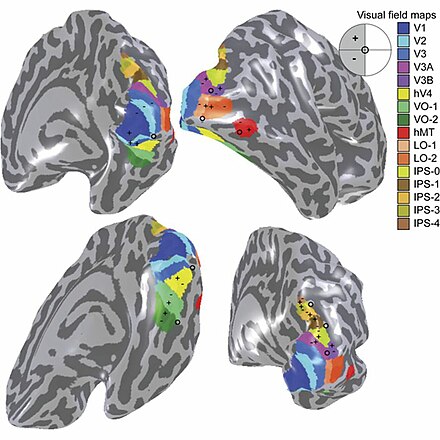
La corteza visual primaria V1 envía información visual a las áreas corticales extraestriadas para un procesamiento visual de orden superior. Estas áreas corticales extraestriadas se ubican por delante del lóbulo occipital. Los principales se designan como áreas visuales V2, V3, V4 y V5 / MT. Cada área puede tener múltiples funciones. Hallazgos recientes han demostrado que el centro de color no está aislado ni es rastreable a un área única en la corteza visual. Más bien, hay múltiples áreas que posiblemente tengan diferentes roles en la capacidad de procesar los estímulos de color.
Área visual V4
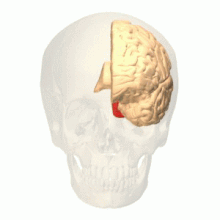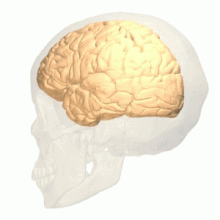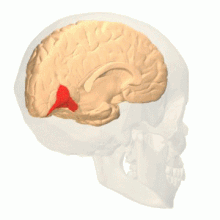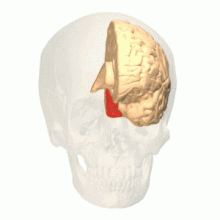

Los estudios anatómicos y fisiológicos han establecido que el centro de color comienza en V1 y envía señales a las áreas extraestriadas V2 y V4 para su posterior procesamiento. V4 en particular es un área de interés debido a la fuerza de los campos receptivos de color en sus neuronas. [4] V4 se identificó inicialmente en experimentos de corteza visual de monos macacos. Originalmente, se propuso que el color se procesara de forma selectiva en V4. Sin embargo, esta hipótesis fue luego rechazada a favor de otra hipótesis que sugería que V4 y otras áreas alrededor de V4 trabajan juntas para procesar el color en forma de múltiples regiones selectivas de color. [5]Después de la identificación de V4 como la región selectiva del color en los monos macacos, los científicos comenzaron a buscar una estructura homóloga en la corteza humana. Utilizando imágenes cerebrales de resonancia magnética funcional, los científicos encontraron tres áreas principales estimuladas por el color: V1, un área en el lóbulo occipital ventral, específicamente la circunvolución lingual, que fue designada como V4 humana, o hV4, y otra área ubicada anteriormente en la circunvolución fusiforme, designada como V4α. [4] [6]
El propósito de V4 ha cambiado dinámicamente a medida que se realizan nuevos estudios. Dado que V4 responde fuertemente al color tanto en monos macacos como en humanos, se ha convertido en un área de interés para los científicos. [6] El área V4 se atribuyó originalmente a la selectividad del color, pero nuevas pruebas han demostrado que V4, así como otras áreas de la corteza visual, son receptivas a diversas entradas. Las neuronas V4 son receptivas a una serie de propiedades, como el color, el brillo y la textura. También participa en el procesamiento de forma, orientación, curvatura, movimiento y profundidad. [7]
La organización real de hV4 en la corteza todavía se está investigando. En el mono macaco, V4 abarca la dorsal y ventral.lóbulo occipital. Los experimentos humanos han demostrado que V4 solo se extiende por la porción ventral. Esto llevó a distinguir el hV4 del macaco V4. Un estudio reciente de Winawer et al. El análisis de las mediciones de fMRI para mapear el hV4 y las áreas occipitales ventrales mostró que las variaciones entre los sujetos utilizados para el mapeo de hV4 se atribuyeron al principio a un error de instrumentación, pero Winawer argumentó que los senos en el cerebro interferían con las mediciones de fMRI. Se probaron dos modelos para hV4: un modelo tenía hV4 completamente en el lado ventral y el segundo modelo tenía hV4 dividido en secciones dorsal y ventral. Se llegó a la conclusión de que aún era difícil trazar un mapa de la actividad de hV4 y que se requería más investigación. Sin embargo, otra evidencia, como lesiones en el lóbulo occipital ventral que causan acromatopsia, sugirió que el área occipital ventral juega un papel importante en la visión del color. [8]
V4α
La búsqueda del equivalente humano de V4 condujo al descubrimiento de otras áreas estimuladas por el color. El más significativo fue un área anterior en el lóbulo occipital ventral, posteriormente denominada V4α. Otros experimentos de resonancia magnética funcional encontraron que V4α tenía una función diferente a la de V4, pero trabajaba de manera cooperativa con ella. [1] V4α está involucrado en una serie de procesos y está activo durante las tareas que requieren el orden de los colores, las imágenes, el conocimiento sobre el color, las ilusiones de color y el color del objeto.
Complejo V4-V4α
Las áreas V4 y V4α son entidades separadas, pero debido a su proximidad en la circunvolución fusiforme, estas dos áreas a menudo se denominan colectivamente el complejo V4. La investigación sobre el complejo V4 descubrió que diferentes estimulaciones cromáticas activaban el área V4 o V4α, y algunos parámetros de estimulación activaban ambos. Por ejemplo, las imágenes de colores naturales activaron V4α con más fuerza que V4. Las imágenes de colores no naturales activaron tanto V4α como V4 por igual. Se concluyó que las dos subdivisiones cooperan entre sí para generar imágenes en color, pero también están funcionalmente separadas. [4]
Un estudio de Nunn et al. sobre la activación del complejo V4 en personas con sinestesia visual al escuchar palabras habladas se utilizó para predecir la ubicación del centro de color. La sinestesia es el fenómeno donde un estímulo sensorial produce una reacción automática e involuntaria en una sensación diferente . En este estudio, se estudiaron las personas que verían colores al escuchar palabras para ver si la reacción del color se podía rastrear hasta un área cortical específica. Los resultados de la resonancia magnética funcional mostraron que la circunvolución fusiforme izquierda, un área compatible con V4, se activaba cuando los sujetos hablaban. También encontraron una activación simultánea de V4α. Hubo poca actividad en las áreas V1 y V2. Estos resultados validaron la existencia del complejo V4 en humanos como un área especializada para la visión del color. [9]
Corteza preestriada V2
Se cree que V2, también llamada corteza preestriada, tiene un pequeño papel en el procesamiento del color al proyectar señales desde V1 al complejo V4. Aún se está investigando si las células selectivas de color están presentes en V2. Algunos estudios de imágenes ópticas han encontrado pequeños grupos de células selectivas de color rojo-verde en V1 y V2, pero no células selectivas de color azul-amarillo. [1] Otros estudios han demostrado que V2 se activa por estímulos de color, pero no color después de imágenes. [8] V4 también tiene retroalimentación sobre V2, lo que sugiere que existe una red definida de comunicación entre las múltiples áreas de la corteza visual. Cuando se inyectó GABA , un neurotransmisor inhibidor , en las células V4, las células V2 experimentaron una disminución significativa de la excitabilidad. [10]
Métodos de investigación
La resonancia magnética funcional , o fMRI para abreviar, ha sido clave para determinar las regiones selectivas de color en la corteza visual. La fMRI puede rastrear la actividad cerebral midiendo el flujo sanguíneo en todo el cerebro. Las áreas que tienen más sangre fluyendo hacia ellas indican una ocurrencia de actividad neuronal. Este cambio en el flujo sanguíneo se denomina respuesta hemodinámica . Entre los beneficios de la fMRI se incluye el mapeo dinámico en tiempo real de los procesos corticales. Sin embargo, la fMRI no puede rastrear la activación real de las neuronas, que ocurre en una escala de tiempo de milisegundos, pero puede rastrear la respuesta hemodinámica, que ocurre en una escala de tiempo de segundos. Este método es ideal para rastrear neuronas selectivas de color porque la percepción del color da como resultado una imagen residual visual.que se puede observar en las neuronas, que dura unos 15 segundos. [11]
Sakai y col. utilizaron fMRI para observar si la activación de la circunvolución fusiforme se correlacionaba con la percepción del color y la imagen posterior. Los sujetos del estudio Sakai se colocaron en la máquina de resonancia magnética funcional y posteriormente se sometieron a diversos estímulos visuales. Se mostró una serie de tres imágenes a los sujetos mientras se usaba fMRI para enfocar la hemodinámica de la circunvolución fusiforme. La primera imagen fue un patrón de seis círculos de colores. Las siguientes dos imágenes fueron acromáticas. Una de las imágenes tenía una cruz gris y la otra imagen tenía los mismos seis círculos que la primera imagen, excepto que eran seis tonos de gris que se correlacionaban con las imágenes coloreadas. Los sujetos fueron ciclados entre el círculo y las imágenes en cruz. Durante las imágenes cruzadas, el sujeto percibió una imagen secundaria.Los resultados del experimento mostraron que hubo un aumento significativo de actividad en la circunvolución fusiforme cuando el sujeto vio la imagen en color. Esto proporcionó más evidencia de la existencia del centro de color fuera de la corteza visual primaria.[11]
Acromatopsia cerebral
La acromatopsia cerebral es una afección crónica en la que una persona no puede ver el color, pero aún puede reconocer la forma y la forma. La acromatopsia cerebral se diferencia de la acromatopsia congénita en que es causada por daño en la corteza cerebral en contraposición a anomalías en las células de la retina. La búsqueda del centro de color fue motivada por el descubrimiento de que las lesiones en el lóbulo occipital ventral conducían al daltonismo, así como por la idea de que existen áreas de especialización en la corteza. Muchos estudios han demostrado que las lesiones en las áreas comúnmente identificadas como el centro de color, como V1, V2 y el complejo V4, conducen a acromatopsia. [1]La acromatopsia cerebral ocurre después de una lesión en la circunvolución lingual o fusiforme, las áreas asociadas con hV4. Estas lesiones incluyen trauma físico, accidente cerebrovascular y crecimiento de tumores. Una de las principales iniciativas para localizar el centro de color en la corteza visual es descubrir la causa y un posible tratamiento de la acromatopsia cerebral.
El alcance de los síntomas y el daño es diferente de una persona a otra. Si una persona tiene acromatopsia completa, todo su campo visual está desprovisto de color. Una persona con discromatopsia, o acromtopsia incompleta, tiene síntomas similares a la acromatopsia completa, pero en menor grado. Esto puede ocurrir en personas que tenían acromatopsia, pero el cerebro se recuperó de la lesión, restaurando algo de la visión de los colores. La persona puede ver ciertos colores. Sin embargo, hay muchos casos en los que no hay recuperación. Finalmente, una persona con hemiacromatopsia ve la mitad de su campo de visión en color y la otra mitad en gris. El hemicampo visual contralateral a una lesión en la circunvolución lingual o fusiforme es el que aparece gris, mientras que el hemicampo visual ipsilateral aparece en color. [11] La variación en los síntomas enfatiza la necesidad de comprender la arquitectura del centro de color para diagnosticar mejor y posiblemente tratar la acromotopsia cerebral.
Referencias
- ↑ a b c d e f Shapley R., Hawken MJ (2011). "Color en la corteza: células oponentes simples y dobles" . Investigación de la visión . 51 (7): 701–717. doi : 10.1016 / j.visres.2011.02.012 . PMC 3121536 . PMID 21333672 .
- ^ Conway BR (15 de abril de 2001). "Estructura espacial de las entradas del cono a las células de color en la corteza visual primaria de macacos alerta (V-1)" . J. Neurosci . 21 (8): 2768–83. doi : 10.1523 / JNEUROSCI.21-08-02768.2001 . PMC 6762533 . PMID 11306629 .
- ^ Livingstone MS, Hubel DH (1984). "Anatomía y fisiología de un sistema de color en la corteza visual de primates" . Revista de neurociencia . 4 : 309–356. doi : 10.1523 / jneurosci.04-01-00309.1984 . PMID 6198495 .
- ↑ a b c Bartels A., Zeki S. (2000). "La arquitectura del centro de color en el cerebro visual humano: nuevos resultados y una revisión". La Revista Europea de Neurociencia . 12 (1): 172-193. doi : 10.1046 / j.1460-9568.2000.00905.x . PMID 10651872 .
- ^ Tootell RBH, Nelissen K., Vanduffel W., Orban GA (2004). "Búsqueda de color 'Centro (s)' en Macaque Visual Cortex" . Corteza cerebral . 14 (4): 353–363. doi : 10.1093 / cercor / bhh001 . PMID 15028640 . CS1 maint: varios nombres: lista de autores ( enlace )
- ↑ a b Murphey DK, Yoshor D., Beauchamp Michael S. (2008). "La percepción coincide con la selectividad en el centro de color anterior humano" . Biología actual . 18 (3): 216–220. doi : 10.1016 / j.cub.2008.01.013 . PMID 18258428 . CS1 maint: varios nombres: lista de autores ( enlace )
- ^ Roe Anna W. (2012). "Hacia una teoría unificada del área visual V4" . Neurona . 74 (1): 12-29. doi : 10.1016 / j.neuron.2012.03.011 .
- ^ Winawer J., Horiguchi H., Sayres RA, Amano K., Wandell BA (2010). "Mapeo de hV4 y corteza occipital ventral: el eclipse venoso" . Revista de visión . 10 : 5. doi : 10.1167 / 10.5.1 .CS1 maint: varios nombres: lista de autores ( enlace )
- ^ Nunn JA, Gregory LJ, Brammer M., Williams SCR, Parslow DM, Morgan MJ, Gray JA (2002). "Imagen de resonancia magnética funcional de la sinestesia: activación de V4 / V8 por palabras habladas. [Artículo]". Neurociencia de la naturaleza . 5 (4): 371–375. doi : 10.1038 / nn818 . PMID 11914723 . CS1 maint: varios nombres: lista de autores ( enlace )
- ^ Jansen-Amorim AK, Fiorani M., Gattass R. (2012). "La inactivación de GABA del área V4 cambia las propiedades del campo receptivo de las neuronas V2 en monos Cebus" . Neurología experimental . 235 (2): 553–562. doi : 10.1016 / j.expneurol.2012.03.008 .CS1 maint: varios nombres: lista de autores ( enlace )
- ↑ a b c Sakai K., Watanabe E., Onodera Y., Uchida I., Kato H., Yamamoto E., Miyashita Y. (1995). "Mapeo funcional del centro de color humano con imágenes de resonancia magnética eco-planar". Actas: Ciencias Biológicas . 261 (1360): 89–98. doi : 10.1098 / rspb.1995.0121 .CS1 maint: varios nombres: lista de autores ( enlace )