Las especies heterotálicas tienen sexos que residen en diferentes individuos. El término se aplica particularmente para distinguir los hongos heterotálicos , que requieren dos parejas compatibles para producir esporas sexuales, de los homotálicos , que son capaces de reproducirse sexualmente a partir de un solo organismo.
En los hongos heterotálicos, dos individuos diferentes aportan núcleos para formar un cigoto. Se incluyen ejemplos de heterotalismo para Saccharomyces cerevisiae , Aspergillus fumigatus , Aspergillus flavus , Penicillium marneffei y Neurospora crassa . El ciclo de vida heterotálico de N. crassa se da con cierto detalle, ya que existen ciclos de vida similares en otros hongos heterotálicos.
Ciclo de vida de Saccharomyces cerevisiae
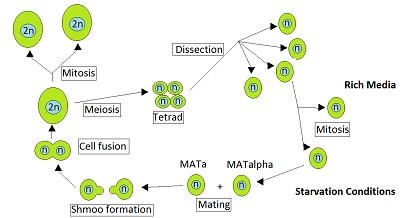
La levadura Saccharomyces cerevisiae es heterotálica. Esto significa que cada célula de levadura es de un cierto tipo de apareamiento y solo puede aparearse con una célula del otro tipo de apareamiento. Durante el crecimiento vegetativo que normalmente ocurre cuando los nutrientes son abundantes, S. cerevisiae se reproduce por mitosis como células haploides o diploides. Sin embargo, cuando mueren de hambre, las células diploides se someten a meiosis para formar esporas haploides. [1] El apareamiento ocurre cuando las células haploides del tipo de apareamiento opuesto, MATa y MATα, entran en contacto. Ruderfer y col. [2] señaló que tales contactos son frecuentes entre células de levadura estrechamente relacionadas por dos razones. La primera es que las células del tipo de apareamiento opuesto están presentes juntas en el mismo ascus., el saco que contiene la tétrada de células producidas directamente por una sola meiosis , y estas células pueden aparearse entre sí. La segunda razón es que las células haploides de un tipo de apareamiento, tras la división celular, a menudo producen células del tipo de apareamiento opuesto con las que pueden aparearse.
Katz Ezov y col. [3] presentó evidencia de que en las poblaciones naturales de S. cerevisiae predominan la reproducción clonal y un tipo de “autofertilización” (en forma de apareamiento intratetrado). Ruderfer y col. [2] analizó la ascendencia de cepas naturales de S. cerevisiae y concluyó que el cruzamiento se produce solo una vez cada 50.000 divisiones celulares. Por tanto, aunque S. cerevisiae es heterotálico, parece que, en la naturaleza, el apareamiento se produce con mayor frecuencia entre células de levadura estrechamente relacionadas. La relativa rareza en la naturaleza de los eventos meióticos que resultan del cruzamiento exterior sugiere que los posibles beneficios a largo plazo del cruzamiento exterior (p. Ej., Generación de diversidad genética) es poco probable que sean suficientes para mantener el sexo de una generación a la siguiente. [ cita requerida ] Más bien, un beneficio a corto plazo, como la reparación recombinacional meiótica de los daños en el ADN causados por condiciones estresantes como la inanición, puede ser la clave para el mantenimiento del sexo en S. cerevisiae . [4] [5]
Ciclo de vida de Aspergillus fumigatus
Aspergillus fumigatus , es un hongo heterotálico. [6] Es una de lasespecies de Aspergillus más comunesque causa enfermedades en humanos con inmunodeficiencia . A. fumigatus , está muy extendido en la naturaleza y se encuentra típicamente en el suelo y la materia orgánica en descomposición, como los montones de compost, donde juega un papel esencial en el reciclaje de carbono y nitrógeno . Las colonias del hongo producen a partir de conidióforos miles de conidios diminutos de color verde grisáceo(2-3 μm) que se transportan fácilmente por el aire. A. fumigatus posee un ciclo reproductivo sexual completamente funcional que conduce a la producción de cleistotecia yascosporas . [7]
Aunque A. fumigatus se encuentra en áreas con climas y ambientes muy diferentes, muestra una baja variación genética y falta de diferenciación genética poblacional a escala global. [8] Por lo tanto, la capacidad para el sexo heterotálico se mantiene a pesar de que se produce poca diversidad genética. Como en el caso anterior de S. cereviae , un beneficio a corto plazo de la meiosis puede ser la clave para el mantenimiento adaptativo del sexo en esta especie.
Ciclo de vida de Aspergillus flavus
A. flavus es el principal productor de aflatoxinas cancerígenas en los cultivos de todo el mundo. También es un patógeno oportunista para humanos y animales , que causa aspergilosis en individuos inmunodeprimidos. En 2009, se descubrió que surgía un estado sexual de este hongo heterotálico cuando se cultivaban juntas cepas del tipo de apareamiento opuesto en condiciones adecuadas. [9]
La sexualidad genera diversidad en el grupo de genes de aflatoxinas en A. flavus , [10] lo que sugiere que la producción de variación genética puede contribuir al mantenimiento del heterotalismo en esta especie.
Ciclo de vida de Talaromyces marneffei
Henk y col. [11] mostró que los genes necesarios para la meiosis están presentes en T. marneffei, y que el apareamiento y la recombinación genética ocurren en esta especie.
Henk y col. [11] concluyó que T. marneffei se reproduce sexualmente, pero la recombinación en poblaciones naturales es más probable que ocurra a través de distancias espacial y genéticamente limitadas, lo que resulta en una estructura poblacional altamente clonal. El sexo se mantiene en esta especie aunque se produce muy poca variabilidad genética . El sexo puede mantenerse en T. marneffei por un beneficio a corto plazo de la meiosis, como en S. cerevisiae y A. fumigatus , ya comentadas.
Ciclo de vida de Neurospora crassa

El ciclo sexual de N. crassa es heterotálico. Los cuerpos de fructificación sexual (peritecios) solo se pueden formar cuando se unen dos micelios de diferente tipo de apareamiento. Como otros ascomicetos , N. crassa tiene dos tipos de apareamiento que, en este caso, están simbolizados por 'A' y 'a'. No hay una diferencia morfológica evidente entre las cepas de tipo 'A' y de tipo apareamiento. Ambos pueden formar abundante protoperitecia, la estructura reproductora femenina (ver figura, parte superior de § ). Los protoperitecios se forman más fácilmente en el laboratorio cuando el crecimiento ocurre en un medio sintético sólido (agar) con una fuente relativamente baja de nitrógeno. [12] La inanición de nitrógeno parece ser necesaria para la expresión de genes implicados en el desarrollo sexual.[13] El protoperitecio consiste en un ascogonio, una hifa multicelular enrollada que está encerrada en una agregación de hifas en forma de nudo. Un sistema ramificado de hifas delgadas, llamado tricogyne, se extiende desde la punta del ascogonio y se proyecta más allá de las hifas envolventes en el aire. El ciclo sexual se inicia (es decir, se produce la fertilización) cuando una célula (normalmente un conidio) de tipo de apareamiento opuesto entra en contacto con una parte del tricogyne (ver figura, parte superior de §). Dicho contacto puede ir seguido de la fusión celular que conduce a que uno o más núcleos de la célula fertilizante migren por el tricoginio hacia el ascogonio. Dado que las cepas 'A' y 'a' tienen las mismas estructuras sexuales, ninguna de las cepas puede considerarse exclusivamente masculina o femenina. Sin embargo, como receptor, el protoperitecio de las cepas 'A' y 'a' puede considerarse la estructura femenina, y el conidio fertilizante puede considerarse el participante masculino.
Los pasos subsiguientes que siguen a la fusión de las células haploides 'A' y 'a' han sido delineados por Fincham y Day, [14] y por Wagner y Mitchell. [15]Después de la fusión de las células, se retrasa la fusión adicional de sus núcleos. En cambio, un núcleo de la célula fertilizante y un núcleo del ascogonio se asocian y comienzan a dividirse sincrónicamente. Los productos de estas divisiones nucleares (todavía en pares de tipos de apareamiento diferentes, es decir, 'A' / 'a') migran a numerosas hifas ascógenas, que luego comienzan a crecer fuera del ascogonio. Cada una de estas hifas ascógenas se dobla para formar un gancho (o báculo) en su punta y el par 'A' y 'a' de núcleos haploides dentro del báculo se dividen sincrónicamente. A continuación, se forman septos para dividir el báculo en tres celdas. La celda central en la curva del gancho contiene un núcleo 'A' y un núcleo 'a' (ver figura, parte superior de § ). Esta célula binuclear inicia la formación de ascus y se denomina célula "inicial de ascus". Siguiente los doslas células uninucleadas a ambos lados de la primera célula formadora de ascus se fusionan entre sí para formar una célula binucleada que puede crecer para formar un báculo adicional que luego puede formar su propia célula inicial de ascus. Este proceso se puede repetir varias veces.
Después de la formación de la célula inicial de ascus, el núcleo 'A' y 'a' se fusionan entre sí para formar un núcleo diploide (ver figura, parte superior de § ). Este núcleo es el único núcleo diploide en todo el ciclo de vida de N. crassa . El núcleo diploide tiene 14 cromosomas formados a partir de dos núcleos haploides fusionados que tenían 7 cromosomas cada uno. La formación del núcleo diploide es seguida inmediatamente por la meiosis . Las dos divisiones secuenciales de la meiosis conducen a cuatro núcleos haploides, dos del tipo de apareamiento 'A' y dos del tipo de apareamiento 'a'. Una división mitótica adicional conduce a cuatro núcleos 'A' y cuatro núcleos 'a' en cada ascus . Mitosises una parte esencial del ciclo de vida de todos los organismos que se reproducen sexualmente y, en sus características principales, la meiosis en N. crassa parece típica de la meiosis en general.
A medida que ocurren los eventos anteriores, la vaina micelial que había envuelto el ascogonio se desarrolla como la pared del peritecio, se impregna de melanina y se ennegrece. El peritecio maduro tiene una estructura en forma de matraz.
Un peritecio maduro puede contener hasta 300 asci, cada uno derivado de núcleos diploides de fusión idénticos. Normalmente, en la naturaleza, cuando los peritecios maduran, las ascosporas son expulsadas con bastante violencia al aire. Estas ascosporas son resistentes al calor y, en el laboratorio, requieren calentamiento a 60 ° C durante 30 minutos para inducir la germinación. Para las cepas normales, el ciclo sexual completo toma de 10 a 15 días. En un asco maduro que contiene 8 ascosporas, los pares de esporas adyacentes son idénticos en constitución genética, ya que la última división es mitótica, y dado que las ascosporas están contenidas en el saco de ascos que las mantiene en un orden definido determinado por la dirección de las segregaciones nucleares durante mitosis. Dado que los cuatro productos primarios también están ordenados en secuencia,el patrón de marcadores genéticos de una segregación de primera división se puede distinguir de los marcadores de un patrón de segregación de segunda división.
Ver también
- Apareamiento de levadura
Referencias
- ^ Herskowitz I (diciembre de 1988). "Ciclo de vida de la levadura en ciernes Saccharomyces cerevisiae" . Microbiol. Rev . 52 (4): 536–53. doi : 10.1128 / MMBR.52.4.536-553.1988 . PMC 373162 . PMID 3070323 .
- ↑ a b Ruderfer DM, Pratt SC, Seidel HS, Kruglyak L (septiembre de 2006). "Análisis genómico poblacional de cruzamiento y recombinación en levadura" . Nat. Genet . 38 (9): 1077–81. doi : 10.1038 / ng1859 . PMID 16892060 .
- ^ Katz Ezov T, Chang SL, Frenkel Z, Segrè AV, Bahalul M, Murray AW, Leu JY, Korol A, Kashi Y (enero de 2010). "Heterotalismo en Saccharomyces cerevisiae aislados de la naturaleza: efecto del locus HO sobre el modo de reproducción" . Mol. Ecol . 19 (1): 121–31. doi : 10.1111 / j.1365-294X.2009.04436.x . PMC 3892377 . PMID 20002587 .
- ^ Birdsell JA, Testamentos C (2003). El origen evolutivo y el mantenimiento de la recombinación sexual: una revisión de los modelos contemporáneos. Serie de biología evolutiva >> Biología evolutiva, vol. 33 págs. 27-137. MacIntyre, Ross J .; Clegg, Michael, T (Eds.), Springer. ISBN 978-0306472619
- ^ Elvira Hörandl (2013). La meiosis y la paradoja del sexo en la naturaleza, meiosis, ISBN 978-953-51-1197-9 , InTech, DOI: 10.5772 / 56542
- ^ Sugui JA, Losada L, Wang W, Varga J, Ngamskulrungroj P, Abu-Asab M, Chang YC, O'Gorman CM, Wickes BL, Nierman WC, Dyer PS, Kwon-Chung KJ (2011). "Identificación y caracterización de una pareja" supermater Aspergillus fumigatus " . mBio . 2 (6): e00234–11. doi : 10.1128 / mBio.00234-11 . PMC 3225970 . PMID 22108383 .
- ^ O'Gorman CM, Fuller H, Dyer PS (enero de 2009). "Descubrimiento de un ciclo sexual en el hongo patógeno oportunista Aspergillus fumigatus" . Naturaleza . 457 (7228): 471–4. Código bibliográfico : 2009Natur.457..471O . doi : 10.1038 / nature07528 . PMID 19043401 .
- ^ Rydholm C, Szakacs G, Lutzoni F (abril de 2006). "Baja variación genética y ninguna estructura de población detectable en Aspergillus fumigatus en comparación con especies de Neosartorya estrechamente relacionadas" . Célula eucariota . 5 (4): 650–7. doi : 10.1128 / EC.5.4.650-657.2006 . PMC 1459663 . PMID 16607012 .
- ^ Cuerno BW, Moore GG, Carbone I (2009). "Reproducción sexual en Aspergillus flavus" . Micología . 101 (3): 423–9. doi : 10.3852 / 09-011 . PMID 19537215 .
- ^ Moore GG, Elliott JL, Singh R, Horn BW, Dorner JW, Stone EA, Chulze SN, Barros GG, Naik MK, Wright GC, Hell K, Carbone I (2013). "La sexualidad genera diversidad en el grupo de genes de aflatoxinas: evidencia a escala mundial" . PLoS Pathog . 9 (8): e1003574. doi : 10.1371 / journal.ppat.1003574 . PMC 3757046 . PMID 24009506 .
- ^ a b Henk DA, Shahar-Golan R, Devi KR, Boyce KJ, Zhan N, Fedorova ND, Nierman WC, Hsueh PR, Yuen KY, Sieu TP, Kinh NV, Wertheim H, Baker SG, Day JN, Vanittanakom N, Bignell EM, Andrianopoulos A, Fisher MC (2012). "Clonalidad a pesar del sexo: la evolución de los vecindarios sexuales asociados con el anfitrión en el hongo patógeno Penicillium marneffei" . PLoS Pathog . 8 (10): e1002851. doi : 10.1371 / journal.ppat.1002851 . PMC 3464222 . PMID 23055919 .
- ^ Westergaard M, Mitchell HK (1947). "Neurospora. Parte V. Un medio sintético que favorece la reproducción sexual". Revista estadounidense de botánica . 34 : 573–577.
- ^ Nelson MA, Metzenberg RL (septiembre de 1992). "Genes de desarrollo sexual de Neurospora crassa" . Genética . 132 (1): 149-162. PMC 1205113 . PMID 1356883 .
- ^ Fincham J RS, Día PR (1963). Genética de hongos . Oxford, Reino Unido: Blackwell Scientific Publications. ASIN B000W851KO .
- ^ Wagner RP, Mitchell HK (1964). Genética y metabolismo . Nueva York, NY: John Wiley and Sons. ASIN B00BXTC5BO .