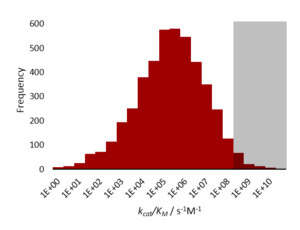
Una enzima de difusión limitada cataliza una reacción de forma tan eficaz que la etapa de limitación de la velocidad es la de difusión del sustrato en el sitio activo o difusión del producto . [2] Esto también se conoce como perfección cinética o perfección catalítica . Dado que la velocidad de catálisis de tales enzimas está determinada por la reacción de difusión controlada , por lo tanto, representa una restricción física intrínseca a la evolución (una altura máxima de pico en el panorama de aptitud física). Las enzimas perfectas de difusión limitada son muy raras. La mayoría de las enzimas catalizan sus reacciones a una velocidad entre 1.000 y 10.000 veces más lenta que este límite. Esto se debe tanto a las limitaciones químicas de las reacciones difíciles como a las limitaciones evolutivas de que velocidades de reacción tan altas no confieren ninguna aptitud adicional . [1]
Historia

La teoría de la reacción controlada por difusión fue utilizada originalmente por RA Alberty, Gordon Hammes y Manfred Eigen para estimar el límite superior de la reacción enzima-sustrato. [3] [4] Según su estimación, [3] [4] el límite superior de la reacción enzima-sustrato fue 10 9 M −1 s −1 .
En 1972, se observó que en la deshidratación de H 2 CO 3 catalizada por anhidrasa carbónica , la constante de velocidad de segundo orden obtenida experimentalmente fue de aproximadamente 1,5 × 10 10 M −1 s −1 , [5] que era un orden de magnitud más alto que el límite superior estimado por Alberty, Hammes y Eigen basado en un modelo simplificado. [3] [4]
Para abordar tal paradoja, Kuo-Chen Chou y sus colaboradores propusieron un modelo teniendo en cuenta el factor espacial y el factor de campo de fuerza entre la enzima y su sustrato y encontraron que el límite superior podría alcanzar 10 10 M −1 s - 1 , [6] [7] [8] y puede usarse para explicar algunas tasas de reacción sorprendentemente altas en biología molecular. [5] [9] [10]
El nuevo límite superior encontrado por Chou et al. para la reacción enzima-sustrato se discutió y analizó más a fondo mediante una serie de estudios de seguimiento. [11] [12] [13]
Se elaboró una comparación detallada entre el modelo simplificado de Alberty-Hammes-Eigen ( a ) y el modelo de Chou ( b ) para calcular la velocidad de reacción controlada por difusión de la enzima con su sustrato, o el límite superior de la reacción enzima-sustrato, se elaboró en la papel. [14]
Mecanismo
Las enzimas cinéticamente perfectas tienen una constante de especificidad , k cat / K m , del orden de 10 8 a 10 9 M −1 s −1 . La velocidad de la reacción catalizada por enzimas está limitada por la difusión, por lo que la enzima "procesa" el sustrato mucho antes de que encuentre otra molécula. [1]
Algunas enzimas operan con cinéticas que son más rápidas que las velocidades de difusión, lo que parecería imposible. Se han invocado varios mecanismos para explicar este fenómeno. Se cree que algunas proteínas aceleran la catálisis atrayendo su sustrato y preorientándolas mediante el uso de campos eléctricos dipolares. Algunos invocan una explicación de efecto túnel mecánico-cuántico mediante la cual un protón o un electrón pueden atravesar barreras de activación. Si la teoría del túnel de protones siguió siendo una idea controvertida, [15] [16] se ha demostrado que es el único mecanismo posible en el caso de la lipoxigenasa de la soja. [17]
Evolución
Vale la pena señalar que no hay muchas enzimas cinéticamente perfectas. Esto se puede explicar en términos de selección natural . Se puede favorecer un aumento de la velocidad catalítica ya que podría conferir alguna ventaja al organismo. Sin embargo, cuando la velocidad catalítica supera la velocidad de difusión (es decir, los sustratos que entran y salen del sitio activo, y también se encuentran con sustratos), no hay más ventaja para aumentar la velocidad aún más. El límite de difusión representa una restricción física absoluta sobre la evolución. [1] Aumentar la velocidad catalítica más allá de la velocidad de difusión no ayudará al organismo de ninguna manera y, por lo tanto, representa un máximo global en un panorama de fitness . Por lo tanto, estas enzimas perfectas deben haber surgido por una mutación aleatoria 'afortunada'que pasó a extenderse, o porque la velocidad más rápida alguna vez fue útil como parte de una reacción diferente en la ascendencia de la enzima. [ cita requerida ]
Ejemplos
- Acetilcolinesterasa
- β-lactamasa
- Catalasa
- Anhídrido carbónico
- Monóxido de carbono deshidrogenasa [18]
- Peroxidasa del citocromo c
- Fumarase
- Superóxido dismutasa
- Triosafosfato isomerasa
Ver también
- Reacción controlada por difusión
- Enzima
- Catálisis enzimática
- La cinética de enzimas
- Ingeniería enzimática
Referencias
- ^ a b c d Bar-Even, Arren; Noor, Elad; Savir, Yonatan; Liebermeister, Wolfram; Davidi, Dan; Tawfik, Dan S; Milo, Ron (2011). "La enzima moderadamente eficiente: tendencias evolutivas y fisicoquímicas que dan forma a los parámetros de la enzima". Bioquímica . 50 (21): 4402–10. doi : 10.1021 / bi2002289 . PMID 21506553 .
- ↑ Berg, Jeremy M .; Tymoczko, JL; Stryer, L. (2006). "Fosforilación oxidativa". Bioquímica (5 ed.). págs. 491–526. ISBN 978-0716787242.
- ↑ a b c Alberty, Robert A .; Hammes, Gordon G. (1958). "Aplicación de la teoría de las reacciones controladas por difusión a la cinética enzimática". Revista de Química Física . 62 (2): 154–9. doi : 10.1021 / j150560a005 .
- ^ a b c Eigen, Manfred; Hammes, Gordon G. (2006). "Pasos elementales en reacciones enzimáticas (estudiadas por espectrometría de relajación)". En Nord, FF (ed.). Avances en enzimología y áreas afines de la biología molecular . págs. 1-38. doi : 10.1002 / 9780470122709.ch1 . ISBN 978-0-470-12270-9. OCLC 777630506 . PMID 14149678 .
- ↑ a b Koenig, Seymour H .; Brown, Rodney D. (1972). "H 2 CO 3 como sustrato para la anhidrasa carbónica en la deshidratación de HCO 3 - " . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 69 (9): 2422–5. Código Bibliográfico : 1972PNAS ... 69.2422K . doi : 10.1073 / pnas.69.9.2422 . JSTOR 61783 . PMC 426955 . PMID 4627028 .
- ^ Chou, Kuo-Chen; Jiang, Shou-Ping (1974). "Estudios sobre la tasa de reacciones de enzimas controladas por difusión. Factor espacial y factor de campo de fuerza". Scientia Sinica . 27 (5): 664–80. PMID 4219062 .
- ^ Chou, Kuo-Chen (1976). "La cinética de la reacción de combinación entre enzima y sustrato". Scientia Sinica . 19 (4): 505-28. PMID 824728 .
- ^ Li, TT; Chou, KC (1976). "Las relaciones cuantitativas entre la velocidad de reacción controlada por difusión y los parámetros característicos en los sistemas de reacción enzima-sustrato. I. Sustratos neutros". Scientia Sinica . 19 (1): 117–36. PMID 1273571 .
- ^ Riggs, Arthur D; Burguesa, Suzanne; Cohn, Melvin (1970). "La interacción represor-operador lac". Revista de Biología Molecular . 53 (3): 401-17. doi : 10.1016 / 0022-2836 (70) 90074-4 . PMID 4924006 .
- ^ Kirschner, Kasper; Gallego, Ernesto; Schuster, Inge; Goodall, David (1971). "Unión cooperativa de dinucleótido de nicotinamida-adenina a gliceraldehído-3-fosfato deshidrogenasa de levadura". Revista de Biología Molecular . 58 (1): 29–50. doi : 10.1016 / 0022-2836 (71) 90230-0 . PMID 4326080 .
- ^ Chou, Kuo Chen; Zhou, Guo Ping (1982). "Papel de la proteína fuera del sitio activo en la reacción de enzimas controlada por difusión". Revista de la Sociedad Química Estadounidense . 104 (5): 1409-1413. doi : 10.1021 / ja00369a043 .
- ^ Payens, TAJ (1983). "¿Por qué las enzimas son tan grandes?". Tendencias en Ciencias Bioquímicas . 8 (2): 46. doi : 10.1016 / 0968-0004 (83) 90382-1 .
- ^ Zhou, Guozhi; Wong, Ming-Tat; Zhou, Guo-Qiang (1983). "Reacciones de enzimas controladas por difusión". Química biofísica . 18 (2): 125–32. doi : 10.1016 / 0301-4622 (83) 85006-6 . PMID 6626685 .
- ^ Zhou, Guo-Qiang; Zhong, Wei-Zhu (1982). "Reacciones de enzimas controladas por difusión". Revista europea de bioquímica . 128 (2–3): 383–7. doi : 10.1111 / j.1432-1033.1982.tb06976.x . PMID 7151785 .
- ↑ García-Viloca, M; Gao, Jiali; Karplus, Martin; Truhlar, Donald G (2004). "Cómo funcionan las enzimas: análisis por la teoría de la tasa moderna y simulaciones por computadora". Ciencia . 303 (5655): 186–95. Código Bibliográfico : 2004Sci ... 303..186G . doi : 10.1126 / science.1088172 . PMID 14716003 . S2CID 17498715 .
- ^ Olsson, Mats HM; Siegbahn, Per EM; Warshel, Arieh (2004). "Simulaciones del efecto de gran isótopo cinético y la dependencia de la temperatura de la transferencia de átomos de hidrógeno en lipoxigenasa". Revista de la Sociedad Química Estadounidense . 126 (9): 2820–8. doi : 10.1021 / ja037233l . PMID 14995199 .
- ^ Jevtic, S; Anders, J (2017). "Un modelo cualitativo de tasa cuántica para la transferencia de hidrógeno en lipoxigenasa de soja". La Revista de Física Química . 147 (11): 114108. arXiv : 1612.03773 . Código bibliográfico : 2017JChPh.147k4108J . doi : 10.1063 / 1.4998941 . PMID 28938801 . S2CID 11202267 .
- ^ Domnik, Lilith; Merrouch, Meriem; Goetzl, Sebastián; Jeoung, Jae-Hun; Léger, Christophe; Dementin, Sébastien; Fourmond, Vincent; Dobbek, Holger (27 de noviembre de 2017). "CODH-IV: una deshidrogenasa de CO captador de CO de alta eficiencia con resistencia al O2" (PDF) . Angewandte Chemie International Edition . 56 (48): 15466-15469. doi : 10.1002 / anie.201709261 . ISSN 1521-3773 . PMID 29024326 .