Corteza cerebral
| Corteza cerebral | |
|---|---|
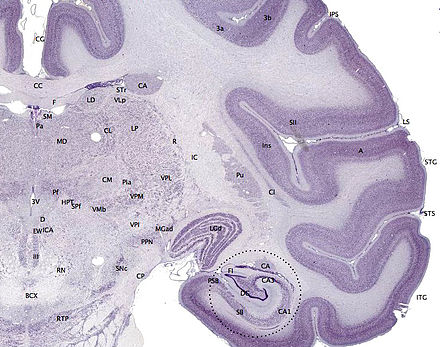 Corte de tejido del cerebro de un mono macaco adulto ( Macaca mulatta ). La corteza cerebral es la capa externa representada en violeta oscuro. Fuente: BrainMaps.org | |
 Neuronas teñidas de Golgi en la corteza | |
| Detalles | |
| Parte de | Cerebro |
| Identificadores | |
| latín | Corteza cerebral |
| Malla | D002540 |
| NeuroNames | 39 |
| Identificación de NeuroLex | birnlex_1494 |
| TA98 | A14.1.09.003 A14.1.09.301 |
| TA2 | 5527 , 5528 |
| FMA | 61830 |
| Términos anatómicos de la neuroanatomía | |
{kind=link}
La corteza cerebral , también conocida como manto cerebral , [1] es la capa externa de tejido neural del cerebro del cerebro en humanos y otros mamíferos . La corteza cerebral consiste principalmente en la neocorteza de seis capas , con solo el 10% de alocorteza . [2] Está separado en dos cortezas , por la fisura longitudinal que divide el cerebro en los hemisferios cerebrales izquierdo y derecho . Los dos hemisferios están unidos debajo de la corteza por el cuerpo calloso.. La corteza cerebral es el sitio más grande de integración neuronal en el sistema nervioso central . [3] Desempeña un papel clave en la atención , la percepción , la conciencia , el pensamiento , la memoria , el lenguaje y la conciencia .
En la mayoría de los mamíferos, aparte de los mamíferos pequeños que tienen cerebros pequeños, la corteza cerebral está plegada, lo que proporciona una mayor superficie en el volumen confinado del cráneo . Además de minimizar el volumen cerebral y craneal, el plegamiento cortical es crucial para el cableado del cerebro y su organización funcional. En los mamíferos con un cerebro pequeño no hay plegamiento y la corteza es lisa. [4] [5]
Un pliegue o cresta en la corteza se denomina circunvolución (plural circunvoluciones) y un surco se denomina surco (plural surcos). Estas circunvoluciones superficiales aparecen durante el desarrollo fetal y continúan madurando después del nacimiento a través del proceso de girificación . En el cerebro humano, la mayor parte de la corteza cerebral no es visible desde el exterior, pero está enterrada en los surcos, [6] y la corteza insular está completamente oculta. Los surcos y circunvoluciones principales marcan las divisiones del cerebro en los lóbulos del cerebro .
Hay entre 14 y 16 mil millones de neuronas en la corteza cerebral humana. Estos están organizados en capas horizontales y radialmente en columnas corticales y minicolumnas . Las áreas corticales tienen funciones específicas como el movimiento en la corteza motora y la vista en la corteza visual .
Estructura
La corteza cerebral es la cubierta exterior de las superficies de los hemisferios cerebrales y se pliega en picos llamados circunvoluciones y surcos llamados surcos . En el cerebro humano tiene entre dos y tres o cuatro milímetros de espesor, [7] y constituye el 40 por ciento de la masa del cerebro. [3] El 90 por ciento de la corteza cerebral es la neocorteza de seis capas y el otro 10 por ciento está formado por alocorteza . [3] Hay entre 14 y 16 mil millones de neuronas en la corteza, [3] y estas están organizadas radialmente en columnas corticales y minicolumnas., en las capas organizadas horizontalmente de la corteza. [8] [9]
La neocorteza se puede separar en diferentes regiones de la corteza conocidas en plural como cortezas, e incluye la corteza motora y la corteza visual . Aproximadamente dos tercios de la superficie cortical están enterrados en los surcos y la corteza insular está completamente oculta. La corteza es más gruesa en la parte superior de una circunvolución y más delgada en la parte inferior de un surco. [10]
Pliegues
La corteza cerebral está plegada de manera que permite que una gran superficie de tejido neural encaje dentro de los límites del neurocráneo . Cuando se despliega en el ser humano, cada corteza hemisférica tiene una superficie total de aproximadamente 0,12 metros cuadrados (1,3 pies cuadrados). [11] El pliegue se produce hacia adentro, alejándose de la superficie del cerebro, y también está presente en la superficie medial de cada hemisferio dentro de la fisura longitudinal . La mayoría de los mamíferos tienen una corteza cerebral que está enredada con los picos conocidos como circunvoluciones y los valles o surcos conocidos como surcos. Algunos pequeños mamíferos, incluidos algunos pequeños roedores, tienen superficies cerebrales lisas sin girificación . [5]
Lóbulos
Los surcos y circunvoluciones más grandes marcan las divisiones de la corteza cerebral en los lóbulos del cerebro . [7] Hay cuatro lóbulos principales: el lóbulo frontal , el lóbulo parietal , el lóbulo temporal y el lóbulo occipital . La corteza insular a menudo se incluye como lóbulo insular. [12] El lóbulo límbico es un borde de la corteza en el lado medial de cada hemisferio y también se incluye a menudo. [13] También se describen tres lóbulos del cerebro: el lóbulo paracentral , el lóbulo parietal superior y el lóbulo parietal inferior..
Espesor
Para las especies de mamíferos, los cerebros más grandes (en términos absolutos, no solo en relación con el tamaño del cuerpo) tienden a tener cortezas más gruesas. [14] Los mamíferos más pequeños, como las musarañas , tienen un grosor neocortical de aproximadamente 0,5 mm; los que tienen el cerebro más grande, como los humanos y las ballenas de aleta, tienen un grosor de 2 a 4 mm. [3] [7] Existe una relación aproximadamente logarítmica entre el peso del cerebro y el grosor cortical. [14] La resonancia magnética del cerebro (IRM) permite obtener una medida del grosor de la corteza cerebral humana y relacionarla con otras medidas. El grosor de las diferentes áreas corticales varía pero, en general, la corteza sensorial es más delgada que la corteza motora.[15] Un estudio ha encontrado alguna asociación positiva entre el grosor cortical y la inteligencia . [16] Otro estudio encontró que la corteza somatosensorial es más gruesa en quienes padecen migraña , aunque no se sabe si esto es el resultado de los ataques de migraña o la causa de ellos. [17] [18] Un estudio posterior que utilizó una población de pacientes más grande no informa ningún cambio en el grosor cortical en pacientes con migraña. [19] Un trastorno genético de la corteza cerebral, por el cual la disminución del plegamiento en ciertas áreas da como resultado un microgiro , donde hay cuatro capas en lugar de seis, en algunos casos se considera que está relacionado con la dislexia .[20]
Capas de neocorteza
El neocórtex está formado por seis capas, numeradas del I al VI, desde la capa más externa I, cerca de la piamadre , hasta la capa más interna VI, cerca de la sustancia blanca subyacente . Cada capa cortical tiene una distribución característica de diferentes neuronas y sus conexiones con otras regiones corticales y subcorticales. Existen conexiones directas entre diferentes áreas corticales y conexiones indirectas a través del tálamo.
Uno de los ejemplos más claros de estratificación cortical es la línea de Gennari en la corteza visual primaria . Se trata de una banda de tejido más blanco que se puede observar a simple vista en el surco calcarino del lóbulo occipital. La línea de Gennari está compuesta por axones que llevan información visual desde el tálamo a la capa IV de la corteza visual .
La tinción de secciones transversales de la corteza para revelar la posición de los cuerpos de las células neuronales y los tractos axónicos intracorticales permitió a los neuroanatomistas a principios del siglo XX producir una descripción detallada de la estructura laminar de la corteza en diferentes especies. El trabajo de Korbinian Brodmann (1909) estableció que el neocórtex de los mamíferos se divide consistentemente en seis capas.
Capa I
La capa I es la capa molecular y contiene pocas neuronas dispersas, incluidas las neuronas GABAérgicas de rosa mosqueta . [21] La capa I consiste principalmente en extensiones de mechones dendríticos apicales de neuronas piramidales y axones orientados horizontalmente, así como células gliales . [22] Durante el desarrollo, las células de Cajal-Retzius [23] y las células de la capa granular subpial [24] están presentes en esta capa. Además, aquí se pueden encontrar algunas células estrelladas espinosas . Se cree que las entradas a los mechones apicales son cruciales para la retroalimentacióninteracciones en la corteza cerebral implicadas en el aprendizaje asociativo y la atención. [25] Si bien alguna vez se pensó que la entrada a la capa I provenía de la corteza misma, [26] ahora se sabe que la capa I a través del manto de la corteza cerebral recibe información sustancial de la matriz o de las células del tálamo de tipo M [27] ( en contraste con el núcleo o el tipo C que van a la capa IV). [28]
Capa II
La capa II, la capa granular externa , contiene pequeñas neuronas piramidales y numerosas neuronas estrelladas.
Capa III
La capa III, la capa piramidal externa , contiene predominantemente neuronas piramidales de tamaño pequeño y mediano, así como neuronas no piramidales con axones intracorticales orientados verticalmente; las capas I a III son el objetivo principal de las aferentes corticocorticales interhemisféricas , y la capa III es la fuente principal de las eferentes corticocorticales .
Capa IV
La capa IV, la capa granular interna , contiene diferentes tipos de células estrelladas y piramidales, y es el principal objetivo de las aferentes talamocorticales de las neuronas tálamo tipo C (tipo núcleo) [28] , así como de las aferentes corticocorticales intrahemisféricas. Las capas superiores a la capa IV también se denominan capas supragranulares (capas I-III), mientras que las capas inferiores se denominan capas infragranulares (capas V y VI).
Capa V
La capa V, la capa piramidal interna , contiene grandes neuronas piramidales. Los axones de estos abandonan la corteza y se conectan con estructuras subcorticales, incluidos los ganglios basales . En la corteza motora primaria del lóbulo frontal, la capa V contiene células piramidales gigantes llamadas células de Betz , cuyos axones viajan a través de la cápsula interna , el tallo cerebral y la médula espinal formando el tracto corticoespinal , que es la vía principal para el control motor voluntario. .
Capa VI
La capa VI, la capa polimórfica o multiforme, contiene pocas neuronas piramidales grandes y muchas neuronas multiformes y piramidales en forma de huso pequeñas; la capa VI envía fibras eferentes al tálamo, estableciendo una interconexión recíproca muy precisa entre la corteza y el tálamo. [29] Es decir, las neuronas de la capa VI de una columna cortical se conectan con las neuronas del tálamo que proporcionan información a la misma columna cortical. Estas conexiones son tanto excitadoras como inhibidoras. Las neuronas envían fibras excitadoras a las neuronas en el tálamo y también envían colaterales al núcleo reticular del tálamo que inhiben estas mismas neuronas del tálamo o las adyacentes a ellas. [30]Una teoría es que debido a que la salida inhibitoria se reduce por la entrada colinérgica a la corteza cerebral, esto proporciona al tronco encefálico un "control de ganancia ajustable para la transmisión de entradas lemniscales ". [30]
Columnas
Las capas corticales no se apilan simplemente una sobre otra; existen conexiones características entre diferentes capas y tipos neuronales, que abarcan todo el espesor de la corteza. Estos microcircuitos corticales se agrupan en columnas corticales y minicolumnas . [31] Se ha propuesto que las minicolumnas son las unidades funcionales básicas de la corteza. [32] En 1957, Vernon Mountcastle demostró que las propiedades funcionales de la corteza cambian abruptamente entre puntos lateralmente adyacentes; sin embargo, son continuos en la dirección perpendicular a la superficie. Trabajos posteriores han proporcionado evidencia de la presencia de columnas corticales funcionalmente distintas en la corteza visual (Hubel yWiesel , 1959), [33] corteza auditiva y corteza asociativa.
Las áreas corticales que carecen de una capa IV se denominan agranulares . Las áreas corticales que tienen solo una capa IV rudimentaria se denominan disgranulares. [34] El procesamiento de la información dentro de cada capa está determinado por diferentes dinámicas temporales, teniendo en las capas II / III una oscilación lenta de 2 Hz, mientras que en la capa V tiene una oscilación rápida de 10-15 Hz. [35]
Tipos de corteza
Con base en las diferencias en la organización laminar, la corteza cerebral se puede clasificar en dos tipos, la gran área del neocórtex que tiene seis capas de células y el área mucho más pequeña de alocorteza que tiene tres o cuatro capas: [2]
- La neocorteza también se conoce como isocorteza o neopallium y es la parte de la corteza cerebral madura con seis capas distintas. Ejemplos de áreas neocorticales incluyen la corteza motora primaria granular y la corteza visual primaria estriada . El neocórtex tiene dos subtipos, el isocortex verdadero y el proisocortex, que es una región de transición entre el isocortex y las regiones del periallocortex.
- El alocortex es la parte de la corteza cerebral con tres o cuatro capas y tiene tres subtipos, el paleocortex con tres láminas corticales, el archicortex que tiene cuatro o cinco, y un área de transición adyacente al alocortex , el periallocortex . Ejemplos de alocorteza son la corteza olfativa y el hipocampo .
Existe un área de transición entre la neocorteza y la alocorteza llamada corteza paralímbica , donde se fusionan las capas 2, 3 y 4. Esta área incorpora el proisocortex del neocórtex y el periallocortex del alocortex. Además, la corteza cerebral se puede clasificar en cuatro lóbulos : el lóbulo frontal , el lóbulo temporal , el lóbulo parietal y el lóbulo occipital , nombrados a partir de los huesos suprayacentes del cráneo.
Suministro y drenaje de sangre
El suministro de sangre a la corteza cerebral es parte de la circulación cerebral . Las arterias cerebrales irrigan la sangre que irriga el cerebro. Esta sangre arterial transporta oxígeno, glucosa y otros nutrientes a la corteza. Las venas cerebrales drenan la sangre desoxigenada y los desechos metabólicos, incluido el dióxido de carbono, de regreso al corazón.
Las principales arterias que irrigan la corteza son la arteria cerebral anterior , la arteria cerebral media y la arteria cerebral posterior . La arteria cerebral anterior irriga las porciones anteriores del cerebro, incluida la mayor parte del lóbulo frontal. La arteria cerebral media irriga los lóbulos parietales, los lóbulos temporales y partes de los lóbulos occipitales. La arteria cerebral media se divide en dos ramas para irrigar el hemisferio izquierdo y derecho, donde se ramifican más. La arteria cerebral posterior irriga los lóbulos occipitales.
El círculo de Willis es el principal sistema sanguíneo que se ocupa del suministro de sangre en el cerebro y la corteza cerebral.
Desarrollo
El desarrollo prenatal de la corteza cerebral es un proceso complejo y finamente sintonizado llamado corticogénesis , influenciado por la interacción entre los genes y el medio ambiente. [36]
Tubo neural
La corteza cerebral se desarrolla desde la parte más anterior, la región del prosencéfalo, del tubo neural . [37] [38] La placa neural se pliega y se cierra para formar el tubo neural . Desde la cavidad dentro del tubo neural se desarrolla el sistema ventricular y, desde las células neuroepiteliales de sus paredes, las neuronas y la glía del sistema nervioso. La parte más anterior (frontal o craneal) de la placa neural, el prosencéfalo , que es evidente antes de que comience la neurulación , da lugar a los hemisferios cerebrales y la corteza posterior. [39]
Desarrollo de neuronas corticales
Las neuronas corticales se generan dentro de la zona ventricular , junto a los ventrículos . Al principio, esta zona contiene células madre neurales , que hacen la transición a células gliales radiales, células progenitoras, que se dividen para producir células gliales y neuronas. [40]
Glía radial
La corteza cerebral está compuesta por una población heterogénea de células que dan lugar a diferentes tipos celulares. La mayoría de estas células se derivan de la migración radial de la glía que forma los diferentes tipos de células del neocórtex y es un período asociado con un aumento de la neurogénesis . De manera similar, el proceso de neurogénesis regula la laminación para formar las diferentes capas de la corteza. Durante este proceso, hay un aumento en la restricción del destino celular que comienza con los progenitores anteriores que dan lugar a cualquier tipo de célula en la corteza y los progenitores posteriores dan lugar solo a las neuronas.de capas superficiales. Este destino celular diferencial crea una topografía de adentro hacia afuera en la corteza con neuronas más jóvenes en capas superficiales y neuronas más viejas en capas más profundas. Además, las neuronas laminares se detienen en la fase S o G2 para establecer una fina distinción entre las diferentes capas corticales. La diferenciación laminar no se completa por completo hasta después del nacimiento, ya que durante el desarrollo las neuronas laminares siguen siendo sensibles a señales extrínsecas y señales ambientales. [41]
Aunque la mayoría de las células que componen la corteza se derivan localmente de la glía radial, existe un subconjunto de la población de neuronas que migran desde otras regiones. La glía radial da lugar a neuronas que tienen forma piramidal y usan glutamato como neurotransmisor , sin embargo, estas células migratorias aportan neuronas que tienen forma de estrella y usan GABA como su principal neurotransmisor. Estas neuronas GABAérgicas son generadas por células progenitoras en la eminencia ganglionar medial (MGE) que migran tangencialmente a la corteza a través de la zona subventricular . Esta migración de neuronas GABAérgicas es particularmente importante ya que los receptores GABAson excitantes durante el desarrollo. Esta excitación es impulsada principalmente por el flujo de iones de cloruro a través del receptor GABA; sin embargo, en los adultos, las concentraciones de cloruro cambian y provocan un flujo de cloruro hacia adentro que hiperpolariza las neuronas postsinápticas . [42] Las fibras gliales producidas en las primeras divisiones de las células progenitoras están orientadas radialmente, abarcando el grosor de la corteza desde la zona ventricular hasta la superficie pial externa , y proporcionan un andamiaje para la migración de neuronas hacia afuera desde la zona ventricular . [43] [44]
Al nacer, hay muy pocas dendritas presentes en el cuerpo celular de la neurona cortical y el axón no está desarrollado. Durante el primer año de vida, las dendritas aumentan drásticamente en número, de modo que pueden acomodar hasta cien mil conexiones sinápticas con otras neuronas. El axón puede desarrollarse para extenderse mucho desde el cuerpo celular. [45]
División asimétrica
Las primeras divisiones de las células progenitoras son simétricas, lo que duplica el número total de células progenitoras en cada ciclo mitótico . Luego, algunas células progenitoras comienzan a dividirse asimétricamente, produciendo una célula postmitótica que migra a lo largo de las fibras gliales radiales, dejando la zona ventricular , y una célula progenitora, que continúa dividiéndose hasta el final del desarrollo, cuando se diferencia en una célula glial o glial. una célula ependimaria . A medida que se alarga la fase G1 de la mitosis , en lo que se considera un alargamiento selectivo del ciclo celular, las neuronas recién nacidas migran a capas más superficiales de la corteza. [46] Las células hijas que migran se convierten encélulas piramidales de la corteza cerebral. [47] El proceso de desarrollo está ordenado en el tiempo y regulado por cientos de genes y mecanismos reguladores epigenéticos . [48]
Organización de capas
La estructura en capas de la corteza cerebral madura se forma durante el desarrollo. Las primeras neuronas piramidales generado a cabo migrate de la zona ventricular y zona subventricular , junto con Reelin -producir neuronas Cajal-Retzius , desde la presiembra . A continuación, una cohorte de neuronas que migran hacia el centro de la preplaca divide esta capa transitoria en la zona marginal superficial , que se convertirá en la capa I del neocórtex maduro, y la subplaca , [49] formando una capa intermedia llamada placa cortical.. Estas células formarán las capas profundas de la corteza madura, las capas cinco y seis. Las neuronas nacidas más tarde migran radialmente hacia la placa cortical más allá de las neuronas de la capa profunda y se convierten en las capas superiores (de dos a cuatro). Por lo tanto, las capas de la corteza se crean en orden de adentro hacia afuera. [50] La única excepción a esta secuencia de neurogénesis de adentro hacia afuera ocurre en la capa I de los primates , en los cuales, a diferencia de los roedores , la neurogénesis continúa durante todo el período de corticogénesis . [51]
Patrones corticales
El mapa de áreas corticales funcionales, que incluyen la corteza visual y motora primaria, se origina a partir de un " protomapa ", [52] que está regulado por señales moleculares como el factor de crecimiento de fibroblastos FGF8 en las primeras etapas del desarrollo embrionario. [53] [54] Estas señales regulan el tamaño, la forma y la posición de las áreas corticales en la superficie del primordio cortical, en parte regulando los gradientes de expresión del factor de transcripción , a través de un proceso llamado patrón cortical . Ejemplos de tales factores de transcripción incluyen los genes EMX2 y PAX6 . [55] Juntos, amboslos factores de transcripción forman un gradiente de expresión opuesto. Pax6 se expresa en gran medida en el polo lateral rostral , mientras que Emx2 se expresa en gran medida en el polo caudomedial . El establecimiento de este gradiente es importante para un desarrollo adecuado. Por ejemplo, las mutaciones en Pax6 pueden hacer que los niveles de expresión de Emx2 se expandan fuera de su dominio de expresión normal, lo que finalmente conduciría a una expansión de las áreas normalmente derivadas de la corteza medial caudal, como la corteza visual . Por el contrario, si ocurren mutaciones en Emx2, puede causar que el dominio que expresa Pax6 se expanda y resulte en la cortical frontal y motora.regiones en aumento. Por lo tanto, los investigadores creen que gradientes y centros de señalización similares próximos a la corteza podrían contribuir a la expresión regional de estos factores de transcripción. [42] Dos señales de patrón muy bien estudiadas para la corteza incluyen FGF y ácido retinoico . Si los FGF se expresan incorrectamente en diferentes áreas de la corteza en desarrollo, se interrumpe el patrón cortical . Específicamente, cuando Fgf8 aumenta en el polo anterior , Emx2 se regula a la baja y un caudalse produce un desplazamiento en la región cortical. Esto finalmente provoca una expansión de las regiones rostrales. Por lo tanto, Fgf8 y otros FGF juegan un papel en la regulación de la expresión de Emx2 y Pax6 y representan cómo la corteza cerebral puede especializarse para diferentes funciones. [42]
La rápida expansión de la superficie cortical está regulada por la cantidad de autorrenovación de las células gliales radiales y está parcialmente regulada por los genes FGF y Notch . [56] Durante el período de neurogénesis cortical y formación de capas, muchos mamíferos superiores comienzan el proceso de girificación , que genera los pliegues característicos de la corteza cerebral. [57] [58] La girificación está regulada por una proteína Trnp1 asociada al ADN [59] y por la señalización de FGF y SHH [60] [61]
Evolución
De todas las diferentes regiones del cerebro, la corteza cerebral muestra la variación evolutiva más grande y ha evolucionado más recientemente. [5] En contraste con los circuitos altamente conservados del bulbo raquídeo , por ejemplo, que cumple funciones críticas como la regulación del ritmo cardíaco y respiratorio, muchas áreas de la corteza cerebral no son estrictamente necesarias para la supervivencia. Por lo tanto, la evolución de la corteza cerebral ha visto el advenimiento y la modificación de nuevas áreas funcionales, en particular las áreas de asociación que no reciben información directamente desde fuera de la corteza. [5]
Una teoría clave de la evolución cortical está incorporada en la hipótesis de la unidad radial y la hipótesis del protomapa relacionada , propuesta por primera vez por Rakic. [62] Esta teoría establece que se forman nuevas áreas corticales mediante la adición de nuevas unidades radiales, que se logra a nivel de células madre . La hipótesis del protomapa establece que la identidad celular y molecular y las características de las neuronas en cada área cortical están especificadas por células madre corticales , conocidas como células gliales radiales , en un mapa primordial. Este mapa está controlado por señalización secretadas proteínas y aguas abajo factores de transcripción . [63] [64] [65]
Función
Conexiones
La corteza cerebral está conectada a varias estructuras subcorticales como el tálamo y los ganglios basales , enviándoles información a través de conexiones eferentes y recibiendo información de ellas a través de conexiones aferentes . La mayor parte de la información sensorial se envía a la corteza cerebral a través del tálamo. Sin embargo, la información olfativa pasa a través del bulbo olfatorio hasta la corteza olfativa ( corteza piriforme ). La mayoría de las conexiones son de un área de la corteza a otra, en lugar de áreas subcorticales; Braitenbergy Schüz (1998) afirman que en las áreas sensoriales primarias, en el nivel cortical donde terminan las fibras de entrada, hasta el 20% de las sinapsis son suministradas por aferentes extracorticales, pero que en otras áreas y otras capas es probable que el porcentaje sea mucho menor. . [66]
Zonas corticales
La totalidad de la corteza cerebral se dividió en 52 áreas diferentes en una presentación temprana de Korbinian Brodmann . Estas áreas, conocidas como áreas de Brodmann , se basan en su citoarquitectura pero también se relacionan con diversas funciones. Un ejemplo es el área 17 de Brodmann, que es la corteza visual primaria .
En términos más generales, la corteza se describe típicamente como compuesta por tres partes: áreas sensoriales, motoras y de asociación.
Áreas sensoriales
Las áreas sensoriales son las áreas corticales que reciben y procesan información de los sentidos . Las partes de la corteza que reciben información sensorial del tálamo se denominan áreas sensoriales primarias. Los sentidos de la vista, el oído y el tacto son atendidos por la corteza visual primaria, la corteza auditiva primaria y la corteza somatosensorial primaria, respectivamente. En general, los dos hemisferios reciben información del lado opuesto (contralateral) del cuerpo . Por ejemplo, la corteza somatosensorial primaria derecha recibe información de las extremidades izquierdas y la corteza visual derecha recibe información del campo visual izquierdo.. La organización de los mapas sensoriales en la corteza refleja la del órgano sensor correspondiente, en lo que se conoce como mapa topográfico . Los puntos vecinos en la corteza visual primaria , por ejemplo, corresponden a puntos vecinos en la retina . Este mapa topográfico se llama mapa retinotópico . De la misma manera, existe un mapa tonotópico en la corteza auditiva primaria y un mapa somatotópico en la corteza sensorial primaria. Este último mapa topográfico del cuerpo en la circunvolución central posterior se ha ilustrado como una representación humana deformada, el homúnculo somatosensorial., donde el tamaño de las diferentes partes del cuerpo refleja la densidad relativa de su inervación. Las áreas con mucha inervación sensorial, como las yemas de los dedos y los labios, requieren más área cortical para procesar una sensación más fina.
Áreas motoras
Las áreas motoras están ubicadas en ambos hemisferios de la corteza. Las áreas motoras están muy relacionadas con el control de los movimientos voluntarios, especialmente los movimientos fragmentados finos realizados por la mano. La mitad derecha del área del motor controla el lado izquierdo del cuerpo y viceversa.
Dos áreas de la corteza se conocen comúnmente como motoras:
- Corteza motora primaria , que ejecuta movimientos voluntarios [ cita requerida ]
- Áreas motoras complementarias y corteza premotora , que seleccionan los movimientos voluntarios. [ cita requerida ]
Además, se han descrito funciones motoras para:
- Corteza parietal posterior , que guía los movimientos voluntarios en el espacio.
- Corteza prefrontal dorsolateral , que decide qué movimientos voluntarios realizar de acuerdo con instrucciones, reglas y pensamientos autogenerados de orden superior.
Justo debajo de la corteza cerebral hay masas subcorticales interconectadas de materia gris llamadas ganglios basales (o núcleos). Los ganglios basales reciben información de la sustancia negra del mesencéfalo y las áreas motoras de la corteza cerebral, y envían señales a ambos lugares. Están involucrados en el control del motor. Se encuentran laterales al tálamo. Los principales componentes de los ganglios basales son el núcleo caudado , el putamen , el globo pálido , la sustancia negra , el núcleo accumbens y el núcleo subtalámico . El putamen y el globo pálido también se conocen colectivamente como núcleo lentiforme., porque juntos forman un cuerpo en forma de lente. El putamen y el núcleo caudado también se denominan colectivamente cuerpo estriado por su apariencia rayada. [67] [68]
Áreas de asociación
Las áreas de asociación son las partes de la corteza cerebral que no pertenecen a las regiones primarias. Funcionan para producir una experiencia perceptiva significativa del mundo, nos permiten interactuar de manera efectiva y respaldan el pensamiento y el lenguaje abstractos. Los lóbulos parietal , temporal y occipital , todos ubicados en la parte posterior de la corteza, integran la información sensorial y la información almacenada en la memoria. El lóbulo frontal o complejo de asociación prefrontal está involucrado en la planificación de acciones y movimientos, así como en el pensamiento abstracto. Globalmente, las áreas de asociación están organizadas como redes distribuidas. [69]Cada red conecta áreas distribuidas en regiones muy espaciadas de la corteza. Las redes distintas se colocan adyacentes entre sí, lo que produce una serie compleja de redes entrelazadas. La organización específica de las redes de asociación se debate con evidencia de interacciones, relaciones jerárquicas y competencia entre redes. [70]
En los seres humanos, las redes de asociación son particularmente importantes para la función del lenguaje. En el pasado se teorizó que las habilidades del lenguaje se localizan en el área de Broca en áreas de la circunvolución frontal inferior izquierda , BA44 y BA45 , para la expresión del lenguaje y en el área de Wernicke BA22 , para la recepción del lenguaje. Sin embargo, se ha demostrado que los procesos de expresión y recepción del lenguaje ocurren en áreas distintas de las estructuras alrededor del surco lateral , incluido el lóbulo frontal, los ganglios basales , el cerebelo y la protuberancia . [71]
Significación clínica
Enfermedades neurodegenerativas tales como la enfermedad de Alzheimer y la enfermedad de Lafora , muestran como un marcador, una atrofia de la materia gris de la corteza cerebral. [73]
Otras enfermedades del sistema nervioso central incluyen trastornos neurológicos como epilepsia , trastornos del movimiento y dificultades en el habla ( afasia ).
El daño cerebral por enfermedad o trauma puede involucrar daño a un lóbulo específico, como en el trastorno del lóbulo frontal , y las funciones asociadas se verán afectadas. La barrera hematoencefálica que sirve para proteger al cerebro de infecciones puede verse comprometida permitiendo la entrada de patógenos .
El feto en desarrollo es susceptible a una variedad de factores ambientales que pueden causar defectos de nacimiento y problemas en el desarrollo posterior. El consumo materno de alcohol, por ejemplo, puede provocar un trastorno del espectro alcohólico fetal . [74] Otros factores que pueden causar trastornos del desarrollo neurológico son tóxicos tales como medicamentos , y la exposición a la radiación a partir de los rayos X . Las infecciones también pueden afectar el desarrollo de la corteza. Una infección viral es una de las causas de la lisencefalia , que da como resultado una corteza lisa sin girificación .
Un tipo de electrocorticografía llamado mapeo de estimulación cortical es un procedimiento invasivo que implica colocar electrodos directamente en el cerebro expuesto para localizar las funciones de áreas específicas de la corteza. Se utiliza en aplicaciones clínicas y terapéuticas, incluido el mapeo prequirúrgico. [75]
Genes asociados con trastornos corticales
Existe una serie de mutaciones genéticas que pueden causar una amplia gama de trastornos genéticos de la corteza cerebral, que incluyen microcefalia , esquizencefalia y tipos de lisencefalia . [76] Las anomalías cromosómicas también pueden provocar una serie de trastornos del desarrollo neurológico , como el síndrome de X frágil y el síndrome de Rett .
MCPH1 codifica microcefalina y los trastornos en esta y en ASPM se asocian con microcefalia. [76] Las mutaciones en el gen NBS1 que codifica la nibrina pueden causar el síndrome de rotura de Nijmegen , caracterizado por microcefalia. [76]
Las mutaciones en EMX2 , [77] y COL4A1 se asocian con esquizencefalia , [78] una condición caracterizada por la ausencia de grandes partes de los hemisferios cerebrales.
Historia
En 1909, Korbinian Brodmann distinguió diferentes áreas del neocórtex basándose en diferencias citoarquitectónicas y dividió la corteza cerebral en 52 regiones. [79]
Rafael Lorente de Nó , alumno de Santiago Ramon y Cajal identificó más de 40 tipos diferentes de neuronas corticales en función de la distribución de sus dendritas y axones. [79]
Otros animales
La corteza cerebral se deriva del palio , una estructura en capas que se encuentra en el prosencéfalo de todos los vertebrados . La forma básica del palio es una capa cilíndrica que encierra ventrículos llenos de líquido. Alrededor de la circunferencia del cilindro hay cuatro zonas, el palio dorsal, el palio medial, el palio ventral y el palio lateral, que se cree que son homólogas a la neocorteza , el hipocampo , la amígdala y la corteza olfatoria , respectivamente.
Hasta hace poco, no se había reconocido en los invertebrados ninguna contraparte de la corteza cerebral. Sin embargo, un estudio publicado en la revista Cell en 2010, basado en perfiles de expresión génica, informó fuertes afinidades entre la corteza cerebral y los cuerpos en forma de hongo del gusano de trapo Platynereis dumerilii . [80] Los cuerpos de los hongos son estructuras en el cerebro de muchos tipos de gusanos y artrópodos que se sabe que juegan un papel importante en el aprendizaje y la memoria; la evidencia genética indica un origen evolutivo común y, por lo tanto, indica que los orígenes de los primeros precursores de la corteza cerebral se remontan a la era precámbrica temprana.
Imágenes Adicionales
Superficie lateral de la corteza cerebral humana.
Superficie medial de la corteza cerebral humana.
Ver también
- Interfaz cerebro-computadora
- Displasia cortical
- Homúnculo cortical
- Mapa cortical
- Corteza elocuente
- EMX1
- Heterotopía de la materia gris
- Sistema límbico
- Lista de regiones del cerebro humano
Referencias
- ^ "manto cerebral" . TheFreeDictionary.com .
- ↑ a b Strominger, Norman L .; Demarest, Robert J .; Laemle, Lois B. (2012). "Corteza cerebral". Sistema nervioso humano de Noback, séptima edición . Prensa Humana. págs. 429–451. doi : 10.1007 / 978-1-61779-779-8_25 . ISBN 978-1-61779-778-1.
- ↑ a b c d e Saladin, Kenneth (2011). Anatomía humana (3ª ed.). McGraw-Hill. págs. 416–422. ISBN 9780071222075.
- ↑ Fernández, V; Llinares-Benadero, C; Borrell, V (17 de mayo de 2016). "Expansión y plegamiento de la corteza cerebral: ¿qué hemos aprendido?" . El diario EMBO . 35 (10): 1021–44. doi : 10.15252 / embj.201593701 . PMC 4868950 . PMID 27056680 .
- ↑ a b c d Rakic, P (octubre de 2009). "Evolución de la neocorteza: una perspectiva desde la biología del desarrollo" . Nature Reviews Neurociencia . 10 (10): 724–35. doi : 10.1038 / nrn2719 . PMC 2913577 . PMID 19763105 .
- ^ Principios de la ciencia neuronal (4ª ed.). McGraw-Hill, División de Profesiones de la Salud. 2000-01-05. ISBN 978-0838577011.
- ↑ a b c Roberts, P (1992). Neuroanatomía (3ª ed.). Springer-Verlag. págs. 86–92. ISBN 9780387977775.
- ^ Lodato, Simona; Arlotta, Paola (13 de noviembre de 2015). "Generando diversidad neuronal en la corteza cerebral de mamíferos" . Revisión anual de biología celular y del desarrollo . 31 (1): 699–720. doi : 10.1146 / annurev-cellbio-100814-125353 . PMC 4778709 . PMID 26359774 .
Las columnas funcionales fueron definidas por primera vez en la corteza por Mountcastle (1957), quien propuso la hipótesis columnar, que establece que la corteza está compuesta por columnas modulares discretas de neuronas, caracterizadas por un perfil de conectividad consistente.
- ^ Ansen-Wilson, LJ; Lipinski, RJ (enero de 2017). "Interacciones gen-ambiente en el desarrollo y disfunción de la interneurona cortical: una revisión de estudios preclínicos" . Neurotoxicología . 58 : 120-129. doi : 10.1016 / j.neuro.2016.12.002 . PMC 5328258 . PMID 27932026 .
- ^ Carpintero (1985). Texto básico de neuroanatomía (3ª ed.). Williams y Wilkins. págs. 348–358. ISBN 978-0683014556.
- ^ Toro, Roberto; Perron, Michel; Pike, Bruce; Más rico, Louis; Veillette, Suzanne; Pausova, Zdenka; Paus, Tomáš (1 de octubre de 2008). "Tamaño del cerebro y plegamiento de la corteza cerebral humana" . Corteza cerebral . 18 (10): 2352–2357. doi : 10.1093 / cercor / bhm261 . ISSN 1047-3211 . PMID 18267953 .
- ^ Nieuwenhuys, R (2012). "La corteza insular: una revisión". Progresos en la investigación del cerebro . 195 : 123–63. doi : 10.1016 / B978-0-444-53860-4.00007-6 . PMID 22230626 .
- ^ Tortora, G; Derrickson, B (2011). Principios de anatomía y fisiología (13ª ed.). Wiley. pag. 549. ISBN 9780470646083.
- ↑ a b Nieuwenhuys R, Donkelaar HJ, Nicholson C (1998). El sistema nervioso central de los vertebrados, Volumen 1 . Saltador. págs. 2011–2012. ISBN 978-3-540-56013-5.
- ^ Frithjof Kruggel; Martina K. Brückner; Thomas Arendt; Christopher J. Wiggins; D. Yves von Cramon (2003). "Analizando la estructura fina neocortical" . Análisis de imágenes médicas . 7 (3): 251–264. doi : 10.1016 / S1361-8415 (03) 00006-9 . hdl : 11858 / 00-001M-0000-0010-9C60-3 . PMID 12946467 .
- ^ Katherine L. Narr; Roger P. Woods; Paul M. Thompson; Philip Szeszko; Dilbert Robinson; Teodora Dimtcheva; Mala Gurbani; Arthur W. Toga; Robert M. Bilder (2007). "Relaciones entre el coeficiente intelectual y el grosor de la materia gris cortical regional en adultos sanos" . Corteza cerebral . 17 (9): 2163–2171. doi : 10.1093 / cercor / bhl125 . PMID 17118969 .
- ^ Alexandre FM DaSilva; Cristina Granziera; Josh Snyder; Nouchine Hadjikhani (2007). "Engrosamiento en la corteza somatosensorial de pacientes con migraña" . Neurología . 69 (21): 1990–1995. doi : 10.1212 / 01.wnl.0000291618.32247.2d . PMC 3757544 . PMID 18025393 .
- ↑ Catharine Paddock (20 de noviembre de 2007). "Las víctimas de migraña tienen una corteza cerebral más gruesa" . Noticias médicas hoy . Archivado desde el original el 11 de mayo de 2008.
- ^ Datte R, Detre JA, et al. (Octubre de 2011). "Ausencia de cambios en el espesor cortical en pacientes con migraña" . Cefalea . 31 (14): 1452–8. doi : 10.1177 / 0333102411421025 . PMC 3512201 . PMID 21911412 .
- ^ Habib M (2000). "La base neurológica de la dislexia del desarrollo: una visión general y una hipótesis de trabajo" . Cerebro . 123 (12): 2373–99. doi : 10.1093 / cerebro / 123.12.2373 . PMID 11099442 .
- ^ "Los científicos identifican un nuevo tipo de célula del cerebro humano" . Instituto Allen . 27 de agosto de 2018.
- ↑ Shipp, Stewart (17 de junio de 2007). "Estructura y función de la corteza cerebral" . Biología actual . 17 (12): R443–9. doi : 10.1016 / j.cub.2007.03.044 . PMC 1870400 . PMID 17580069 .
- ^ Meyer, Gundela; Goffinet, André M .; Fairén, Alfonso (1999). "Artículo principal: ¿Qué es una célula de Cajal-Retzius? Una reevaluación de un tipo de célula clásica basada en observaciones recientes en el neocórtex en desarrollo" . Corteza cerebral . 9 (8): 765–775. doi : 10.1093 / cercor / 9.8.765 . PMID 10600995 .
- ^ Judaš, Miloš; Pletikos, Mihovil (2010). "El descubrimiento de la capa granular subpial en la corteza cerebral humana". Neurociencia traslacional . 1 (3): 255–260. doi : 10.2478 / v10134-010-0037-4 . S2CID 143409890 .
- ^ CD de Gilbert, Sigman M (2007). "Estados cerebrales: influencias de arriba hacia abajo en el procesamiento sensorial" . Neurona . 54 (5): 677–96. doi : 10.1016 / j.neuron.2007.05.019 . PMID 17553419 .
- ^ Cauller L (1995). "Capa I de la neocorteza sensorial primaria: donde de arriba hacia abajo converge de abajo hacia arriba". Behav Brain Res . 71 (1–2): 163–70. doi : 10.1016 / 0166-4328 (95) 00032-1 . PMID 8747184 . S2CID 4015532 .
- ^ Rubio-Garrido P, Pérez-de-Manzo F, Porrero C, Galazo MJ, Clascá F (2009). "La entrada talámica a las dendritas apicales distales en la capa neocortical 1 es masiva y altamente convergente" . Cereb Cortex . 19 (10): 2380–95. doi : 10.1093 / cercor / bhn259 . PMID 19188274 .
- ↑ a b Jones EG (1998). "Punto de vista: el núcleo y la matriz de la organización talámica". Neurociencia . 85 (2): 331–45. doi : 10.1016 / S0306-4522 (97) 00581-2 . PMID 9622234 . S2CID 17846130 .
- ^ Creutzfeldt, O. 1995. Cortex Cerebri . Springer-Verlag.
- ↑ a b Lam YW, Sherman SM (2010). "Organización funcional de la retroalimentación somatosensorial de la capa 6 cortical al tálamo" . Cereb Cortex . 20 (1): 13-24. doi : 10.1093 / cercor / bhp077 . PMC 2792186 . PMID 19447861 .
- ^ Suzuki, IK; Hirata, T (enero de 2013). "La neurogénesis neocortical no es realmente" neo ": un nuevo modelo evolutivo derivado de un estudio comparativo del desarrollo palial del pollo" (PDF) . Desarrollo, crecimiento y diferenciación . 55 (1): 173–87. doi : 10.1111 / dgd.12020 . PMID 23230908 . S2CID 36706690 .
- ^ Mountcastle V (1997). "La organización columnar del neocórtex" . Cerebro . 120 (4): 701–722. doi : 10.1093 / cerebro / 120.4.701 . PMID 9153131 .
- ^ Hubel DH, Wiesel TN (octubre de 1959). "Campos receptivos de neuronas individuales en la corteza estriada del gato" . La revista de fisiología . 148 (3): 574–91. doi : 10.1113 / jphysiol.1959.sp006308 . PMC 1363130 . PMID 14403679 .
- ^ SM Dombrowski, CC Hilgetag y H. Barbas. La arquitectura cuantitativa distingue los sistemas corticales prefrontales en el mono Rhesus Archivado el29 de agosto de 2008en la Wayback Machine .Cereb. Cortex 11: 975–988. "... o carecen (agranular) o tienen sólo una capa granular IV rudimentaria (disgranular)".
- ^ Sun W, Dan Y (2009). "Oscilación de red de capa específica y campo receptivo espacio-temporal en la corteza visual" . Proc Natl Acad Sci USA . 106 (42): 17986–17991. Código Bibliográfico : 2009PNAS..10617986S . doi : 10.1073 / pnas.0903962106 . PMC 2764922 . PMID 19805197 .
- ^ Pletikos, Mihovil; Sousa, Andre MM; et al. (22 de enero de 2014). "Especificación temporal y bilateralidad de la expresión génica topográfica neocortical humana" . Neurona . 81 (2): 321–332. doi : 10.1016 / j.neuron.2013.11.018 . PMC 3931000 . PMID 24373884 .
- ^ Wolpert, Lewis (2015). Principios de desarrollo (Quinta ed.). Reino Unido: Oxford University Press. pag. 533. ISBN 9780199678143.
- ^ Warren N, Caric D, Pratt T, Clausen JA, Asavaritikrai P, Mason JO, Hill RE, Price DJ (1999). "El factor de transcripción, Pax6, es necesario para la proliferación y diferenciación celular en la corteza cerebral en desarrollo" . Corteza cerebral . 9 (6): 627–35. doi : 10.1093 / cercor / 9.6.627 . PMID 10498281 .
- ^ Larsen, W J. Embriología humana 3.ª edición 2001. pp 421-422 ISBN 0-443-06583-7
- ^ Stephen C. Noctor; Alexander C. Flint; Tamily A. Weissman ; Ryan S. Dammerman y Arnold R. Kriegstein (2001). "Las neuronas derivadas de las células gliales radiales establecen unidades radiales en la neocorteza". Naturaleza . 409 (6821): 714–720. Código bibliográfico : 2001Natur.409..714N . doi : 10.1038 / 35055553 . PMID 11217860 . S2CID 3041502 .
- ^ Sur, Mriganka; Leamey, Catherine A. (2001). "Desarrollo y plasticidad de áreas y redes corticales". Nature Reviews Neurociencia . 2 (4): 251–262. doi : 10.1038 / 35067562 . PMID 11283748 . S2CID 893478 .
- ^ a b c Sanes, Dan H .; Reh, Thomas A .; Harris, William A. (2012). Desarrollo del sistema nervioso . ISBN de Elsevier Inc. 978-0-12-374539-2.
- ^ Rakic, P (octubre de 2009). "Evolución de la neocorteza: una perspectiva desde la biología del desarrollo" . Nature Reviews Neurociencia . 10 (10): 724–35. doi : 10.1038 / nrn2719 . PMC 2913577 . PMID 19763105 .
- ^ Rakic, P (noviembre de 1972). "Determinantes citológicos extrínsecos de cesta y patrón dendrítico de células estrelladas en la capa molecular cerebelosa". La Revista de Neurología Comparada . 146 (3): 335–54. doi : 10.1002 / cne.901460304 . PMID 4628749 . S2CID 31900267 .
- ^ Gilbert, Scott (2006). Biología del desarrollo (8ª ed.). Editores Sinauer Associates. págs. 394–395. ISBN 9780878932504.
- ^ Calegari, F; Haubensack W; Haffner C; Huttner WB (2005). "Alargamiento selectivo del ciclo celular en la subpoblación neurogénica de células progenitoras neurales durante el desarrollo del cerebro de ratón" . La Revista de Neurociencia . 25 (28): 6533–8. doi : 10.1523 / jneurosci.0778-05.2005 . PMC 6725437 . PMID 16014714 .
- ^ P. Rakic (1988). "Especificación de áreas corticales cerebrales". Ciencia . 241 (4862): 170-176. Código Bibliográfico : 1988Sci ... 241..170R . doi : 10.1126 / science.3291116 . PMID 3291116 .
- ^ Hu, XL; Wang, Y .; Shen, Q. (2012). "Control epigenético sobre la elección del destino celular en células madre neurales" . Proteínas y células . 3 (4): 278–290. doi : 10.1007 / s13238-012-2916-6 . PMC 4729703 . PMID 22549586 .
- ^ Kostović, Ivica (1990). "Historia del desarrollo de la zona transitoria de la subplaca en la corteza visual y somatosensorial del mono macaco y el cerebro humano". Revista de Neurología Comparada . 297 (3): 441–470. doi : 10.1002 / cne.902970309 . PMID 2398142 . S2CID 21371568 .
- ^ Rakic, P (1 de febrero de 1974). "Neuronas en la corteza visual del mono rhesus: relación sistemática entre el tiempo de origen y la disposición final". Ciencia . 183 (4123): 425–7. Código Bibliográfico : 1974Sci ... 183..425R . doi : 10.1126 / science.183.4123.425 . PMID 4203022 . S2CID 10881759 .
- ^ Zecevic N, Rakic P (2001). "Desarrollo de neuronas de capa I en la corteza cerebral de primates" . La Revista de Neurociencia . 21 (15): 5607-19. doi : 10.1523 / JNEUROSCI.21-15-05607.2001 . PMC 6762645 . PMID 11466432 .
- ^ Rakic, P (8 de julio de 1988). "Especificación de áreas corticales cerebrales". Ciencia . 241 (4862): 170–6. Código Bibliográfico : 1988Sci ... 241..170R . doi : 10.1126 / science.3291116 . PMID 3291116 .
- ↑ Fukuchi-Shimogori, T; Grove, EA (2 de noviembre de 2001). "Patrón de neocorteza por la molécula de señalización secretada FGF8". Ciencia . 294 (5544): 1071–4. Código Bibliográfico : 2001Sci ... 294.1071F . doi : 10.1126 / science.1064252 . PMID 11567107 . S2CID 14807054 .
- ^ Garel, S; Huffman, KJ; Rubenstein, JL (mayo de 2003). "La regionalización molecular de la neocorteza se interrumpe en mutantes hipomórficos Fgf8" . Desarrollo . 130 (9): 1903–14. doi : 10.1242 / dev.00416 . PMID 12642494 .
- ^ Obispo, KM; Goudreau, G; O'Leary, DD (14 de abril de 2000). "Regulación de la identidad del área en la neocorteza de mamíferos por Emx2 y Pax6". Ciencia . 288 (5464): 344–9. Código Bibliográfico : 2000Sci ... 288..344B . doi : 10.1126 / science.288.5464.344 . PMID 10764649 .
- ^ Erupción, BG; Lim, HD; Breunig, JJ; Vaccarino, FM (26 de octubre de 2011). "La señalización de FGF expande el área de superficie cortical embrionaria regulando la neurogénesis dependiente de Notch" . La Revista de Neurociencia . 31 (43): 15604–17. doi : 10.1523 / jneurosci.4439-11.2011 . PMC 3235689 . PMID 22031906 .
- ↑ Rajagopalan, V; Scott, J; Habas, PA; Kim, K; Corbett-Detig, J; Rousseau, F; Barkovich, AJ; Glenn, OA; Studholme, C (23 de febrero de 2011). "Patrones de crecimiento de tejido local subyacentes a la girificación del cerebro humano fetal normal cuantificados en el útero" . La Revista de Neurociencia . 31 (8): 2878–87. doi : 10.1523 / jneurosci.5458-10.2011 . PMC 3093305 . PMID 21414909 .
- ↑ Lui, Jan H .; Hansen, David V .; Kriegstein, Arnold R. (8 de julio de 2011). "Desarrollo y evolución de la neocorteza humana" . Celular . 146 (1): 18–36. doi : 10.1016 / j.cell.2011.06.030 . ISSN 1097-4172 . PMC 3610574 . PMID 21729779 .
- ^ Stahl, Ronny; Walcher, Tessa; De Juan Romero, Camino; Pilz, Gregor Alexander; Cappello, Silvia; Irmler, Martin; Sanz-Aquela, José Miguel; Beckers, Johannes; Blum, Robert (25 de abril de 2013). "Trnp1 regula la expansión y el plegamiento de la corteza cerebral de los mamíferos mediante el control del destino glial radial" . Celular . 153 (3): 535–549. doi : 10.1016 / j.cell.2013.03.027 . ISSN 1097-4172 . PMID 23622239 .
- ^ Wang, Lei; Hou, Shirui; Han, Young-Goo (23 de mayo de 2016). "La señalización de Hedgehog promueve la expansión del progenitor basal y el crecimiento y plegamiento de la neocorteza" . Neurociencia de la naturaleza . 19 (7): 888–96. doi : 10.1038 / nn.4307 . ISSN 1546-1726 . PMC 4925239 . PMID 27214567 .
- ^ Erupción, Brian G .; Tomasi, Simone; Lim, H. David; Suh, Carol Y .; Vaccarino, Flora M. (26 de junio de 2013). "Gyrification cortical inducida por factor de crecimiento de fibroblastos 2 en el cerebro de ratón" . La Revista de Neurociencia . 33 (26): 10802–10814. doi : 10.1523 / JNEUROSCI.3621-12.2013 . ISSN 1529-2401 . PMC 3693057 . PMID 23804101 .
- ^ Rakic, P (8 de julio de 1988). "Especificación de áreas corticales cerebrales". Ciencia . 241 (4862): 170–6. Código Bibliográfico : 1988Sci ... 241..170R . doi : 10.1126 / science.3291116 . PMID 3291116 .
- ↑ Fukuchi-Shimogori, T; Grove, EA (2 de noviembre de 2001). "Patrón de neocorteza por la molécula de señalización secretada FGF8". Ciencia . 294 (5544): 1071–4. Código Bibliográfico : 2001Sci ... 294.1071F . doi : 10.1126 / science.1064252 . PMID 11567107 . S2CID 14807054 .
- ^ Obispo, KM; Goudreau, G; O'Leary, DD (14 de abril de 2000). "Regulación de la identidad del área en la neocorteza de mamíferos por Emx2 y Pax6". Ciencia . 288 (5464): 344–9. Código Bibliográfico : 2000Sci ... 288..344B . doi : 10.1126 / science.288.5464.344 . PMID 10764649 .
- ^ Grove, EA; Fukuchi-Shimogori, T (2003). "Generación del mapa de áreas corticales cerebrales". Revisión anual de neurociencia . 26 : 355–80. doi : 10.1146 / annurev.neuro.26.041002.131137 . PMID 14527269 . S2CID 12282525 .
- ^ Braitenberg, V y Schüz, A 1998. "Cortex: Estadística y geometría de la conectividad neuronal. Segunda edición revisada a fondo" Nueva York: Springer-Verlag
- ^ Saladino, Kenneth. Anatomía y fisiología: la unidad de forma y función, 5th Ed. Nueva York: McGraw-Hill Companies Inc., 2010. Impresión.
- ^ Diccionario médico de Dorland para consumidores de salud, 2008.
- ^ Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, Roffman JL, Smoller JW, Zöllei L, Polimeni JR, Fischl B, Liu H, Buckner RL (2011). "La organización de la corteza cerebral humana estimada por conectividad funcional intrínseca" . Revista de neurofisiología . 106 (3): 1125-1165. doi : 10.1152 / jn.00338.2011 . PMC 3174820 . PMID 21653723 .
- ^ Rupesh Kumar Srivastava; Jürgen Schmidhuber (2014). "Comprensión de las redes competitivas a nivel local". arXiv : 1410,1165 [ cs.NE ].
- ^ Cathy J. Price (2000). "La anatomía del lenguaje: contribuciones de la neuroimagen funcional" . Revista de anatomía . 197 (3): 335–359. doi : 10.1046 / j.1469-7580.2000.19730335.x . PMC 1468137 . PMID 11117622 .
- ^ Kentar, Modar; Mann, Martina; Sahm, Felix; Olivares-Rivera, Arturo; Sánchez-Porras, Renan; Zerelles, Roland; Sakowitz, Oliver W .; Unterberg, Andreas W .; Santos, Edgar (15 de enero de 2020). "Detección de propagación de despolarizaciones en un modelo de oclusión de arteria cerebral media en porcinos". Acta Neurochirurgica . 162 (3): 581–592. doi : 10.1007 / s00701-019-04132-8 . ISSN 0942-0940 . PMID 31940093 . S2CID 210196036 .
- ^ Ortolano S, Vieitez I, et al. (2014). "La pérdida de neuronas corticales subyace en la neuropatología de la enfermedad de Lafora" . Mol Brain . 7 : 7. doi : 10.1186 / 1756-6606-7-7 . PMC 3917365 . PMID 24472629 .
- ^ Mukherjee, Raja AS; Hollins, Sheila (2006). "Trastorno del espectro alcohólico fetal: una visión general" . Revista de la Real Sociedad de Medicina . 99 (6): 298-302. doi : 10.1258 / jrsm.99.6.298 . PMC 1472723 . PMID 16738372 .
- ^ Tarapore, PE; et al. (Agosto 2012). "Mapeo motor multimodal preoperatorio: una comparación de imágenes de magnetoencefalografía, estimulación magnética transcraneal navegada y estimulación cortical directa" . Revista de neurocirugía . 117 (2): 354–62. doi : 10.3171 / 2012.5.JNS112124 . PMC 4060619 . PMID 22702484 .
- ^ a b c Walsh, Christopher A .; Mochida, Ganeshwaran H. (1 de mayo de 2004). "Base genética de las malformaciones del desarrollo de la corteza cerebral" . Archivos de Neurología . 61 (5): 637–640. doi : 10.1001 / archneur.61.5.637 . PMID 15148137 .
- ^ "Homeobox 2 espiráculos vacíos EMX2 [Homo sapiens (humano)] - Gene - NCBI" . www.ncbi.nlm.nih.gov .
- ^ Smigiel, R; Cábala, M; Jakubiak, A; Kodera, H; Sasiadek, MJ; Matsumoto, N; Sasiadek, MM; Saitsu, H (abril de 2016). "Nueva mutación COL4A1 en un bebé con síndrome dismórfico severo con esquizencefalia, calcificaciones periventriculares y cataratas que se asemejan a una infección congénita". Investigación de defectos de nacimiento. Parte A, Teratología clínica y molecular . 106 (4): 304–7. doi : 10.1002 / bdra.23488 . PMID 26879631 .
- ^ a b Principios de la ciencia neuronal . Kandel, Eric R. (5ª ed.). Nueva York. 2013. págs. 347–348. ISBN 9780071390118. OCLC 795553723 .CS1 maint: otros ( enlace )
- ^ Tomer, R; Denes, AS; Tessmar-Raible, K; Arendt, D; Tomer R; Denes AS; Tessmar-Raible K; Arendt D (2010). "El perfil por registro de imágenes revela el origen común de los cuerpos de los hongos anélidos y el palio de los vertebrados" . Celular . 142 (5): 800–809. doi : 10.1016 / j.cell.2010.07.043 . PMID 20813265 . S2CID 917306 .
enlaces externos
- hier-20 en NeuroNames
- Imágenes de cortes de cerebro teñidas que incluyen la "corteza cerebral" en el proyecto BrainMaps
- "La corteza visual primaria" , Webvision: artículo completo sobre la estructura y función de la corteza visual primaria.
- "Tipos de células básicas" , Webvision: Imagen de los tipos de células básicas de la corteza cerebral del mono.
- Corteza cerebral - Base de datos centrada en células
- Corteza cerebral