 | |
 | |
| Identificadores | |
|---|---|
Modelo 3D ( JSmol ) |
|
| 3DMet | |
| 1208946 | |
| CHEBI | |
| CHEMBL | |
| DrugBank | |
| Tarjeta de información ECHA | 100.005.149 |
| Número CE |
|
| 108834 | |
| KEGG | |
| Malla | Flavina-adenina + dinucleótido |
PubChem CID | |
| UNII | |
Tablero CompTox ( EPA ) | |
| |
| |
| Propiedades | |
| C 27 H 33 N 9 O 15 P 2 | |
| Masa molar | 785,557 g · mol −1 |
| Apariencia | Cristales vítreos blancos |
| log P | -1,336 |
| Acidez (p K a ) | 1.128 |
| Basicidad (p K b ) | 12.8689 |
Salvo que se indique lo contrario, los datos se proporcionan para materiales en su estado estándar (a 25 ° C [77 ° F], 100 kPa). | |
| Referencias de Infobox | |
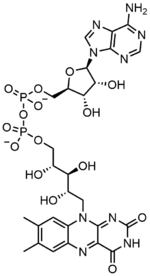
En bioquímica , el dinucleótido de flavina y adenina ( FAD ) es una coenzima con actividad redox asociada con varias proteínas , que participa en varias reacciones enzimáticas del metabolismo . Una flavoproteína es una proteína que contiene un grupo flavina , que puede estar en forma de FAD o mononucleótido de flavina (FMN). Se conocen muchas flavoproteínas: componentes del complejo succinato deshidrogenasa, α-cetoglutarato deshidrogenasa y un componente del complejo piruvato deshidrogenasa .
El FAD puede existir en cuatro estados redox, que son el óxido de flavina-N (5) , quinona , semiquinona e hidroquinona . [1] El FAD se convierte entre estos estados aceptando o donando electrones. El FAD, en su forma completamente oxidada, o forma de quinona , acepta dos electrones y dos protones para convertirse en FADH 2 (forma de hidroquinona). La semiquinona (FADH · ) puede formarse por reducción de FAD u oxidación de FADH 2 aceptando o donando un electrón y un protón, respectivamente. Sin embargo, algunas proteínas generan y mantienen una forma superoxidada del cofactor de flavina, el óxido de flavina-N (5). [2] [3]
Historia
Las flavoproteínas se descubrieron por primera vez en 1879 al separar los componentes de la leche de vaca. Inicialmente se les llamó lactocromo debido a su origen lechoso y pigmento amarillo . [4] La comunidad científica tardó 50 años en lograr un progreso sustancial en la identificación de las moléculas responsables del pigmento amarillo. La década de 1930 lanzó el campo de la investigación de coenzimas con la publicación de muchas estructuras derivadas de flavina y nicotinamida y sus funciones obligatorias en la catálisis redox. Los científicos alemanes Otto Warburg y Walter Christian descubrieron una proteína amarilla derivada de la levadura necesaria para la respiración celularen 1932. Su colega Hugo Theorell separó esta enzima amarilla en apoenzima y pigmento amarillo, y demostró que ni la enzima ni el pigmento eran capaces de oxidar el NADH por sí mismos, pero mezclarlos restauraría la actividad. Theorell confirmó que el pigmento era el éster de fosfato de riboflavina , mononucleótido de flavina (FMN) en 1937, que fue la primera evidencia directa de cofactores enzimáticos . [5] Warburg y Christian encontraron que el FAD era un cofactor de la D-aminoácido oxidasa a través de experimentos similares en 1938. [6]El trabajo de Warburg al vincular la nicotinamida con las transferencias de hidruros y el descubrimiento de las flavinas allanó el camino para que muchos científicos en los años 40 y 50 descubrieran grandes cantidades de bioquímica redox y las unieran en vías como el ciclo del ácido cítrico y la síntesis de ATP .
Propiedades
El dinucleótido de flavina y adenina consta de dos porciones: el nucleótido de adenina ( monofosfato de adenosina ) y el mononucleótido de flavina (FMN) unidos a través de sus grupos fosfato . La adenina se une a una ribosa cíclica en el carbono 1 ' , mientras que el fosfato se une a la ribosa en el carbono 5' para formar el nucleótido de adenina. La riboflavina está formada por un enlace carbono-nitrógeno (CN) entre la isoaloxazina y el ribitol.. El grupo fosfato se une luego al carbono ribosa terminal, formando un FMN. Debido a que el enlace entre la isoaloxazina y el ribitol no se considera un enlace glicosídico , el mononucleótido de flavina no es realmente un nucleótido. [7] Esto hace que el nombre del dinucleótido sea engañoso; sin embargo, el grupo de mononucleótidos de flavina todavía está muy cerca de un nucleótido en su estructura y propiedades químicas.

FAD se puede reducir a FADH 2 mediante la adición de 2 H + y 2 e - . El FADH 2 también se puede oxidar por la pérdida de 1 H + y 1 e - para formar FADH. La forma FAD se puede recrear mediante la pérdida adicional de 1 H + y 1 e - . La formación de FAD también puede ocurrir a través de la reducción y deshidratación del óxido de flavina-N (5). [8] Según el estado de oxidación, las flavinas toman colores específicos cuando están en solución acuosa . flavina-N (5) -óxido(superoxidado) es amarillo anaranjado, FAD (completamente oxidado) es amarillo, FADH (medio reducido) es azul o rojo según el pH , y la forma completamente reducida es incolora. [9] [10] Cambiar la forma puede tener un gran impacto en otras propiedades químicas. Por ejemplo, FAD, la forma completamente oxidada está sujeta a un ataque nucleofílico , la forma completamente reducida, FADH 2 tiene alta polarización , mientras que la forma semi-reducida es inestable en solución acuosa. [11] FAD es un sistema de anillo aromático , mientras que FADH 2 no lo es. [12] Esto significa que FADH 2es significativamente mayor en energía, sin la estabilización por resonancia que proporciona la estructura aromática. El FADH 2 es una molécula portadora de energía, porque una vez oxidada recupera la aromaticidad y libera la energía que representa esta estabilización.
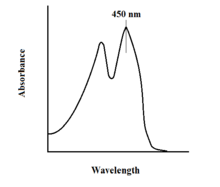
Las propiedades espectroscópicas del FAD y sus variantes permiten el control de la reacción mediante el uso de espectroscopias de fluorescencia y absorción UV-VIS . Cada forma de FAD tiene espectros de absorbancia distintos, lo que facilita la observación de los cambios en el estado de oxidación. [11] Se observa un máximo de absorbancia local importante para FAD a 450 nm, con un coeficiente de extinción de 11.300. [13] Las flavinas en general tienen actividad fluorescente cuando no están unidas (las proteínas unidas a los derivados del ácido nucleico de las flavinas se denominan flavoproteínas ). Esta propiedad se puede utilizar al examinar la unión a proteínas, observando la pérdida de actividad fluorescente cuando se pone en el estado de unión. [11]Las flavinas oxidadas tienen altas absorbancias de aproximadamente 450 nm y emiten fluorescencia a aproximadamente 515-520 nm. [9]
Estados químicos
En los sistemas biológicos, FAD actúa como un aceptor de H + ye - en su forma completamente oxidada, un aceptor o donante en la forma FADH y un donante en la forma FADH 2 reducida . El siguiente diagrama resume los posibles cambios que puede sufrir.

Junto con lo que se ve arriba, se pueden formar y consumir otras formas reactivas de FAD. Estas reacciones implican la transferencia de electrones y la formación / ruptura de enlaces químicos . A través de mecanismos de reacción , FAD puede contribuir a las actividades químicas dentro de los sistemas biológicos. Las siguientes imágenes representan formas generales de algunas de las acciones en las que puede participar FAD.
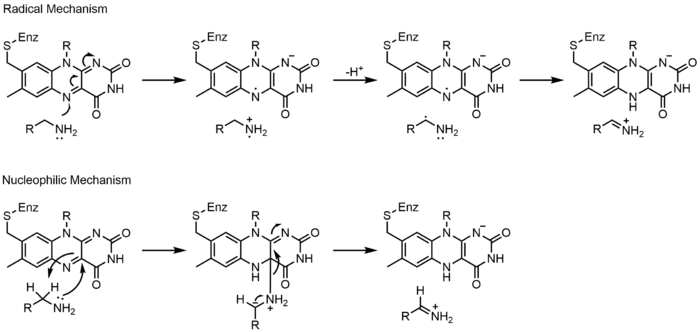
Los mecanismos 1 y 2 representan la ganancia de hidruro , en la que la molécula gana lo que equivale a un ion hidruro. Mecanismos 3 y 4 de formación de radicales y pérdida de hidruros. Las especies radicales contienen átomos de electrones no apareados y son muy activos químicamente. La pérdida de hidruro es el proceso inverso de la ganancia de hidruro visto antes. Los dos últimos mecanismos muestran una adición nucleofílica y una reacción que usa un radical de carbono.
Biosíntesis
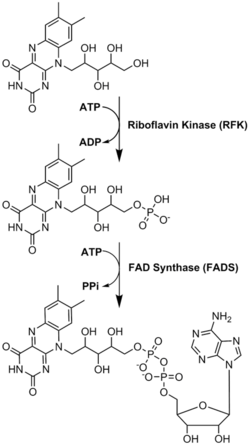
El FAD juega un papel importante como cofactor enzimático junto con el mononucleótido de flavina , otra molécula que se origina a partir de la riboflavina. [8] Las bacterias, los hongos y las plantas pueden producir riboflavina , pero otros eucariotas , como los humanos, han perdido la capacidad de producirla. [9] Por lo tanto, los seres humanos deben obtener riboflavina, también conocida como vitamina B2, de fuentes dietéticas. [14] La riboflavina generalmente se ingiere en el intestino delgado y luego se transporta a las células a través de proteínas transportadoras. [9] La riboflavina quinasa (EC 2.7.1.26) agrega un grupo fosfato a la riboflavina para producir flavina mononucleótido y luego FAD sintetasaune un nucleótido de adenina ; ambos pasos requieren ATP . [9] Las bacterias generalmente tienen una enzima bifuncional, pero las arqueas y los eucariotas suelen emplear dos enzimas distintas. [9] La investigación actual indica que existen distintas isoformas en el citosol y las mitocondrias . [9] Parece que FAD se sintetiza en ambos lugares y potencialmente se transporta donde sea necesario. [11]
Función
Las flavoproteínas utilizan la estructura única y versátil de las fracciones de flavina para catalizar reacciones redox difíciles. Dado que las flavinas tienen múltiples estados redox, pueden participar en procesos que implican la transferencia de uno o dos electrones, átomos de hidrógeno o iones hidronio . El N5 y C4a del anillo de flavina completamente oxidado también son susceptibles al ataque nucleofílico . [15] Esta amplia variedad de ionización y modificación de la fracción flavina se puede atribuir al sistema de anillo de isoaloxazina y la capacidad de las flavoproteínas para perturbar drásticamente los parámetros cinéticos de las flavinas al unirse, incluido el dinucleótido de flavina y adenina (FAD).
El número de genes codificados por proteínas dependientes de flavina en el genoma (el flavoproteoma) depende de la especie y puede oscilar entre el 0,1% y el 3,5%, y los seres humanos tienen 90 genes codificados por flavoproteínas. [16] El FAD es la forma más compleja y abundante de flavina y se informa que se une al 75% del flavoproteoma total [16] y al 84% de las flavoproteínas codificadas en humanos. [17] Se notificaron concentraciones celulares de flavinas libres o unidas no covalentemente en una variedad de líneas de células de mamíferos cultivadas para FAD (2,2-17,0 amol / célula) y FMN (0,46-3,4 amol / célula). [18]
El FAD tiene un potencial de reducción más positivo que el NAD + y es un agente oxidante muy fuerte. La célula utiliza esto en muchas reacciones de oxidación energéticamente difíciles, como la deshidrogenación de un enlace CC a un alqueno . Las proteínas dependientes de FAD funcionan en una gran variedad de vías metabólicas que incluyen transporte de electrones, reparación del ADN, biosíntesis de nucleótidos, beta-oxidación de ácidos grasos, catabolismo de aminoácidos, así como síntesis de otros cofactores como CoA , CoQ y grupos hemo . Una reacción muy conocida es parte del ciclo del ácido cítrico (también conocido como ciclo TCA o Krebs); succinato deshidrogenasa(complejo II en la cadena de transporte de electrones ) requiere FAD unido covalentemente para catalizar la oxidación del succinato a fumarato acoplándolo con la reducción de ubiquinona a ubiquinol . [11] Los electrones de alta energía de esta oxidación se almacenan momentáneamente reduciendo FAD a FADH 2 . FADH 2 luego vuelve a FAD, enviando sus dos electrones de alta energía a través de la cadena de transporte de electrones; la energía en FADH 2 es suficiente para producir 1,5 equivalentes de ATP [19] por fosforilación oxidativa . Algunas flavoproteínas redox se unen de forma no covalente a FAD comoAcetil-CoA-deshidrogenasas que participan en la beta-oxidación de ácidos grasos y el catabolismo de aminoácidos como leucina ( isovaleril-CoA deshidrogenasa ), isoleucina (acil-CoA deshidrogenasa de cadena corta / ramificada), valina (isobutiril-CoA deshidrogenasa) y lisina ( glutaril-CoA deshidrogenasa ). [20] Otros ejemplos de enzimas dependientes de FAD que regulan el metabolismo son la glicerol-3-fosfato deshidrogenasa (síntesis de triglicéridos) y la xantina oxidasa implicadas en el catabolismo de nucleótidos de purina . [21] Las funciones no catalíticas que el FAD puede desempeñar en las flavoproteínas incluyen como roles estructurales, o involucrados en los fotorreceptores de luz sensibles al azul que regulan los relojes biológicos y el desarrollo, la generación de luz en bacterias bioluminiscentes . [20]
Flavoproteínas
Las flavoproteínas tienen una molécula FMN o FAD como grupo protésico, este grupo protésico puede estar unido estrechamente o unido covalentemente. Solo alrededor del 5-10% de las flavoproteínas tienen un FAD unido covalentemente, pero estas enzimas tienen un poder redox más fuerte. [11] En algunos casos, FAD puede proporcionar soporte estructural para sitios activos o proporcionar estabilización de intermedios durante la catálisis. [20] Sobre la base de los datos estructurales disponibles, los sitios de unión a FAD conocidos se pueden dividir en más de 200 tipos. [22]
90 flavoproteínas están codificadas en el genoma humano; alrededor del 84% requiere FAD y alrededor del 16% requiere FMN, mientras que 5 proteínas requieren que ambas estén presentes. [17] Las flavoproteínas se encuentran principalmente en las mitocondrias debido a su poder redox. [17] De todas las flavoproteínas, el 90% realiza reacciones redox y el otro 10% son transferasas , liasas , isomerasas , ligasas . [dieciséis]
Oxidación de enlaces carbono-heteroátomo
Carbono-nitrógeno
La monoamino oxidasa (MAO) es una flavoenzima ampliamente estudiada debido a su importancia biológica con el catabolismo de la noradrenalina , la serotonina y la dopamina . MAO oxida aminas primarias, secundarias y terciarias, que se hidrolizan de forma no enzimática de la imina a aldehído o cetona . Aunque esta clase de enzima se ha estudiado extensamente, su mecanismo de acción todavía se debate. Se han propuesto dos mecanismos: un mecanismo radical y un mecanismo nucleofílico. El mecanismo radical es menos aceptado generalmente porque no hay resonancia paramagnética espectral o electrónica.existe evidencia de la presencia de un intermedio radical. El mecanismo nucleofílico se ve más favorecido porque está respaldado por estudios de mutagénesis dirigida al sitio que mutaron dos residuos de tirosina que se esperaba que aumentaran la nucleofilia de los sustratos. [23]
Carbono-oxígeno
La glucosa oxidasa (GOX) cataliza la oxidación de la β-D-glucosa a D-glucono-δ-lactona con la reducción simultánea de la flavina unida a la enzima. GOX existe como un homodímero, y cada subunidad se une a una molécula de FAD. Las estructuras cristalinas muestran que FAD se une en un bolsillo profundo de la enzima cerca de la interfaz del dímero. Los estudios mostraron que después de la sustitución de FAD con 8-hidroxi-5-carba-5-deaza FAD, la estereoquímica de la reacción se determinó por reacción con el re cara de la flavina. Durante el recambio, se observan las semiquinonas neutras y aniónicas lo que indica un mecanismo radical. [23]

Carbono-azufre
La prenilcisteína liasa (PCLase) cataliza la escisión de prenilcisteína (una modificación de la proteína) para formar un aldehído isoprenoide y el residuo de cisteína liberado en la proteína diana. El FAD está unido de forma no covalente a PCLase. No se han realizado muchos estudios mecanicistas sobre las reacciones de la flavina, pero el mecanismo propuesto se muestra a continuación. Se propone una transferencia de hidruro del C1 del resto prenilo a FAD, lo que da como resultado la reducción de la flavina a FADH 2 . COformED ES un carbocatión que es estabilizado por el átomo de azufre vecino. El FADH 2 luego reacciona con el oxígeno molecular para restaurar la enzima oxidada. [23]

Carbono-carbono
La UDP-N-acetilenolpiruvilglucosamina reductasa (MurB) es una enzima que cataliza la reducción dependiente de NADPH de enolpiruvil-UDP-N-acetilglucosamina (sustrato) al correspondiente compuesto D-lactílico ácido UDP-N-acetilmurámico (producto). MurB es un monómero y contiene una molécula de FAD. Antes de que el sustrato se pueda convertir en producto, NADPH primero debe reducir FAD. Una vez que NADP + se disocia, el sustrato puede unirse y la flavina reducida puede reducir el producto. [23]

Química de tiol / disulfuro
La glutatión reductasa (GR) cataliza la reducción de disulfuro de glutatión (GSSG) a glutatión (GSH). GR requiere FAD y NADPH para facilitar esta reacción; primero se debe transferir un hidruro de NADPH a FAD. La flavina reducida puede actuar entonces como un nucleófilo para atacar el disulfuro, esto forma el aducto C4a-cisteína. La eliminación de este aducto da como resultado un complejo de transferencia de carga de tiolato de flavina. [23]
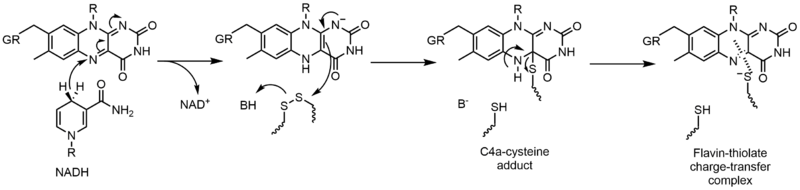
Reacciones de transferencia de electrones
Las enzimas de tipo citocromo P450 que catalizan reacciones de monooxigenasa (hidroxilación) dependen de la transferencia de dos electrones del FAD al P450. En eucariotas se encuentran dos tipos de sistemas P450. Los sistemas P450 que se encuentran en el retículo endoplásmico dependen de una citocromo P-450 reductasa (CPR) que contiene tanto un FAD como un FMN . Los dos electrones en FAD reducido (FADH 2 ) se transfieren uno a la vez al FMN y luego se pasa un solo electrón del FMN al hemo del P450. [24]
Los sistemas P450 que se encuentran en las mitocondrias dependen de dos proteínas de transferencia de electrones: un FAD que contiene adrenodoxina reductasa (AR) y un pequeño grupo de hierro-azufre que contiene una proteína llamada adrenodoxina . FAD está incrustado en el dominio de unión a FAD de AR. [25] [26] El FAD de AR se reduce a FADH 2 mediante la transferencia de dos electrones de NADPH que se unen al dominio de unión a NADP de AR. La estructura de esta enzima está altamente conservada para mantener con precisión la alineación del NADPH donante de electrones y el FAD del aceptor para una transferencia de electrones eficiente. [26]Los dos electrones en FAD reducido se transfieren uno a la vez a la adrenodoxina que a su vez dona el único electrón al grupo hemo del P450 mitocondrial. [27]
Las estructuras de la reductasa del microsoma frente a la reductasa de los sistemas mitocondriales P450 son completamente diferentes y no muestran homología. [24]
Redox
p -hidroxibenzoato hidroxilasa (PHBH) cataliza la oxigenación de p- hidroxibenzoato ( p OHB) a 3,4-dihidroxibenzoato (3,4-diOHB); FAD, NADPH y oxígeno molecular son todos necesarios para esta reacción. NADPH primero transfiere un hidruro equivalente a FAD, creando FADH - , y luego NADP + se disocia de la enzima. El PHBH reducido luego reacciona con el oxígeno molecular para formar el hidroperóxido de flavina-C (4a). El hidroperóxido de flavina hidroxila rápidamente p OHB y luego elimina el agua para regenerar la flavina oxidada. [23] Un mecanismo alternativo de oxigenación mediado por flavina implica el uso de un óxido de flavina-N (5) en lugar de un peróxido de flavina-C (4a) - (hidro).[2] [3]
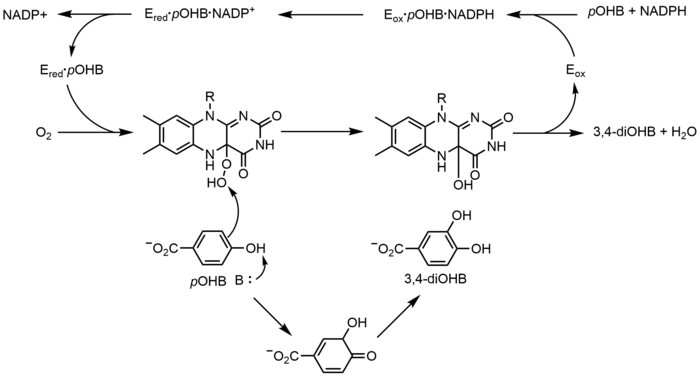
No redox
El corismato sintasa (CS) cataliza el último paso en la vía del shikimato: la formación del corismato. Se conocen dos clases de CS, las cuales requieren FMN , pero se dividen en su necesidad de NADPH como agente reductor. El mecanismo propuesto para CS involucra especies radicales. La especie de radical flavina no se ha detectado espectroscópicamente sin utilizar un análogo de sustrato, lo que sugiere que tiene una vida corta. Sin embargo, al usar un sustrato fluorado, se detectó una semiquinona de flavina neutra. [23]
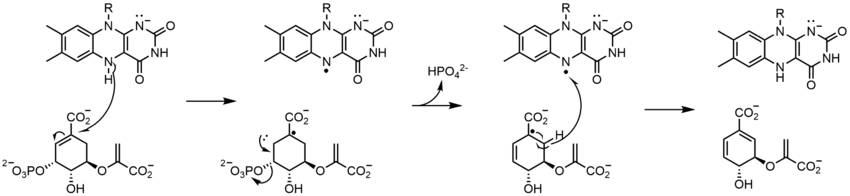
Flavoenzimas complejas
La glutamato sintasa cataliza la conversión de 2-oxoglutarato en L-glutamato con L-glutamina que actúa como fuente de nitrógeno para la reacción. Todas las síntesis de glutamato son flavoproteínas de hierro-azufre que contienen un grupo de hierro-azufre y FMN. Las tres clases de síntesis de glutamato se clasifican en función de sus secuencias y propiedades bioquímicas. Aunque hay tres clases de esta enzima, se cree que todas operan a través del mismo mecanismo, solo difiriendo por lo que primero reduce el FMN. La enzima produce dos moléculas de glutamato: una por hidrólisis de glutamina (formando glutamato y amoniaco) y la segunda por el amoniaco producido en la primera reacción que ataca al 2-oxoglutarato, que es reducido por FMN a glutamato. [23]
Importancia clínica
Debido a la importancia de las flavoproteínas , no es sorprendente que aproximadamente el 60% de las flavoproteínas humanas provoquen enfermedades humanas cuando muten. [17] En algunos casos, esto se debe a una menor afinidad por FAD o FMN, por lo que la ingesta excesiva de riboflavina puede disminuir los síntomas de la enfermedad, como la deficiencia múltiple de acil-CoA deshidrogenasa . [9] Además, la deficiencia de riboflavina en sí misma (y la falta resultante de FAD y FMN) puede causar problemas de salud. [9] Por ejemplo, en pacientes con ELA , hay niveles reducidos de síntesis de FAD. [9] Ambas vías pueden provocar una variedad de síntomas, que incluyen anomalías del desarrollo o gastrointestinales, degradación defectuosa de la grasa , anemia , problemas neurológicos, cáncer o enfermedades cardíacas , migraña , empeoramiento de la visión y lesiones cutáneas. [9] Por tanto, la industria farmacéutica produce riboflavina para complementar la dieta en determinados casos. En 2008, la necesidad mundial de riboflavina fue de 6.000 toneladas por año, con una capacidad de producción de 10.000 toneladas. [4] Este mercado de entre 150 y 500 millones de dólares no es solo para aplicaciones médicas, sino que también se utiliza como complemento de la alimentación animal en la industria agrícola y como colorante alimentario . [4]
Diseño de fármacos
El nuevo diseño de medicamentos antibacterianos es de importancia continua en la investigación científica a medida que aumenta la resistencia de los antibióticos bacterianos a los antibióticos comunes. Una proteína metabólica específica que usa FAD ( Complejo II ) es vital para la virulencia bacteriana, por lo que dirigirse a la síntesis de FAD o crear análogos de FAD podría ser un área útil de investigación. [28] Los científicos ya han determinado las dos estructuras que el FAD suele asumir una vez unido: una conformación extendida o en mariposa, en la que la molécula esencialmente se dobla por la mitad, lo que resulta en el apilamiento de los anillos de adenina e isoaloxazina. [14] Los imitadores de FAD que pueden unirse de manera similar pero que no permiten la función de la proteína podrían ser mecanismos útiles para inhibir la infección bacteriana. [14] Alternativamente, los fármacos que bloquean la síntesis de FAD podrían lograr el mismo objetivo; esto es especialmente intrigante porque la síntesis de FAD humana y bacteriana se basa en enzimas muy diferentes, lo que significa que es poco probable que un fármaco elaborado para atacar la FAD sintasa bacteriana interfiera con las enzimas FAD sintasa humana. [29]
Optogenética
La optogenética permite el control de eventos biológicos de forma no invasiva. [30] El campo ha avanzado en los últimos años con una serie de nuevas herramientas, incluidas las que activan la sensibilidad a la luz, como los dominios FAD que utilizan la luz azul (BLUF). Los BLUF codifican una secuencia de aminoácidos de 100 a 140 que se derivó de fotorreceptores en plantas y bacterias. [30] Al igual que otros fotorreceptores , la luz provoca cambios estructurales en el dominio BLUF que resulta en la interrupción de las interacciones aguas abajo. [30] La investigación actual investiga proteínas con el dominio BLUF adjunto y cómo diferentes factores externos pueden afectar las proteínas. [30]
Seguimiento del tratamiento
Hay una serie de moléculas en el cuerpo que tienen fluorescencia nativa, como triptófano, colágeno , FAD, NADH y porfirinas . [31] Los científicos se han aprovechado de esto al usarlos para monitorear la progresión de la enfermedad o la efectividad del tratamiento o ayudar en el diagnóstico. Por ejemplo, la fluorescencia nativa de un FAD y NADH varía en tejido normal y fibrosis submucosa oral , que es un signo temprano de cáncer oral invasivo . [31] Por lo tanto, los médicos han estado empleando la fluorescencia para ayudar en el diagnóstico y monitorear el tratamiento en lugar de la biopsia estándar . [31]
Imágenes adicionales

Riboflavina
FADH 2


Ver también
- FMN
- FMO , monooxigenasa que contiene flavina
- NAD
Referencias
- ^ Teufel, Robin; Agarwal, Vinayak; Moore, Bradley S. (1 de abril de 2016). "Catálisis de flavoenzimas inusuales en bacterias marinas" . Opinión actual en biología química . 31 : 31–39. doi : 10.1016 / j.cbpa.2016.01.001 . ISSN 1879-0402 . PMC 4870101 . PMID 26803009 .
- ^ a b Teufel, R; Miyanaga, A; Michaudel, Q; Stull, F; Louie, G; Noel, JP; Baran, PS; Palfey, B; Moore, BS (28 de noviembre de 2013). "La oxidación dual mediada por flavina controla un reordenamiento enzimático de tipo Favorskii" . Naturaleza . 503 (7477): 552–6. doi : 10.1038 / nature12643 . PMC 3844076 . PMID 24162851 .
- ^ a b Teufel, Robin; Stull, Frederick; Meehan, Michael J .; Michaudel, Quentin; Dorrestein, Pieter C .; Palfey, Bruce; Moore, Bradley S. (1 de julio de 2015). "Establecimiento bioquímico y caracterización del cofactor Flavin-N5-oxide de EncM" . Revista de la Sociedad Química Estadounidense . 137 (25): 8078–8085. doi : 10.1021 / jacs.5b03983 . ISSN 1520-5126 . PMC 4720136 . PMID 26067765 .
- ↑ a b c Abbas CA, Sibirny AA (junio de 2011). "Control genético de la biosíntesis y transporte de nucleótidos de riboflavina y flavina y construcción de productores biotecnológicos robustos" . Revisiones de Microbiología y Biología Molecular . 75 (2): 321–60. doi : 10.1128 / mmbr.00030-10 . PMC 3122625 . PMID 21646432 .
- ^ Hayashi H (2013). Vitaminas B y folato: química, análisis, función y efectos . Cambridge, Reino Unido: The Royal Society of Chemistry. pag. 7. ISBN 978-1-84973-369-4.
- ^ Warburg O, Christian W (1938). "Aislamiento del grupo protésico del aminoácido oxidasa". Biochemische Zeitschrift . 298 : 150-168.
- ^ Metzler DE, Metzler CM, Sauke DJ (2003). Bioquímica (2ª ed.). San Diego: Harcourt, Academic Press. ISBN 978-0-12-492541-0.
- ↑ a b Devlin TM (2011). Libro de texto de bioquímica: con correlaciones clínicas (7ª ed.). Hoboken, Nueva Jersey: John Wiley & Sons. ISBN 978-0-470-28173-4.
- ↑ a b c d e f g h i j k Barile M, Giancaspero TA, Brizio C, Panebianco C, Indiveri C, Galluccio M, Vergani L, Eberini I, Gianazza E (2013). "Biosíntesis de cofactores flavin en el hombre: implicaciones en la salud y la enfermedad". Diseño Farmacéutico Actual . 19 (14): 2649–75. doi : 10.2174 / 1381612811319140014 . PMID 23116402 .
- ^ Teufel, Robin; Miyanaga, Akimasa; Michaudel, Quentin; Stull, Frederick; Louie, Gordon; Noel, Joseph P .; Baran, Phil S .; Palfey, Bruce; Moore, Bradley S. (28 de noviembre de 2013). "La oxidación dual mediada por flavina controla un reordenamiento enzimático de tipo Favorskii" . Naturaleza . 503 (7477): 552–556. doi : 10.1038 / nature12643 . ISSN 1476-4687 . PMC 3844076 . PMID 24162851 .
- ↑ a b c d e f Kim HJ, Winge DR (mayo de 2013). "Conceptos emergentes en la flavinilación de succinato deshidrogenasa" . Biochimica et Biophysica Acta (BBA) - Bioenergética . 1827 (5): 627–36. doi : 10.1016 / j.bbabio.2013.01.012 . PMC 3626088 . PMID 23380393 .
- ^ Liu S (2012). Ingeniería de Bioprocesos: Cinética, Sostenibilidad y Diseño de Reactores . Newnes. ISBN 978-0-444-63783-3.
- ^ Lewis JA, Escalante-Semerena JC (agosto de 2006). "La enzima tricarballylate deshidrogenasa (TcuA) dependiente de FAD de Salmonella enterica convierte tricarballylate en cis-aconitato" . Revista de bacteriología . 188 (15): 5479–86. doi : 10.1128 / jb.00514-06 . PMC 1540016 . PMID 16855237 .
- ↑ a b c Kuppuraj G, Kruise D, Yura K (noviembre de 2014). "Comportamiento conformacional del dinucleótido de flavina adenina: estereoquímica conservada en estados unidos y libres". El Journal of Physical Chemistry B . 118 (47): 13486–97. doi : 10.1021 / jp507629n . PMID 25389798 .
- ^ Monteira M (2013). Vitaminas B y folato: química, análisis, función y efectos . Cambridge, Reino Unido: The Royal Society of Chemistry. pag. 94. ISBN 978-1-84973-369-4.
- ^ a b c Macheroux P, Kappes B, Ealick SE (agosto de 2011). "Flavogenomics - una vista genómica y estructural de proteínas dependientes de flavina" . La revista FEBS . 278 (15): 2625–34. doi : 10.1111 / j.1742-4658.2011.08202.x . PMID 21635694 . S2CID 22220250 .
- ↑ a b c d Lienhart WD, Gudipati V, Macheroux P (julio de 2013). "El flavoproteoma humano" . Archivos de Bioquímica y Biofísica . 535 (2): 150–62. doi : 10.1016 / j.abb.2013.02.015 . PMC 3684772 . PMID 23500531 .
- ^ Hühner J, Ingles-Prieto Á, Neusüß C, Lämmerhofer M, Janovjak H (febrero de 2015). "Cuantificación de riboflavina, mononucleótido de flavina y dinucleótido de flavina y adenina en células modelo de mamífero por CE con detección de fluorescencia inducida por LED". Electroforesis . 36 (4): 518-25. doi : 10.1002 / elps.201400451 . PMID 25488801 . S2CID 27285540 .
- ^ Stryer L, Berg JM, Tymoczko JL (2007). Bioquímica (6ª ed.). Nueva York: Freeman. ISBN 978-0-7167-8724-2.
- ^ a b c Mansoorabadi SO, Thibodeaux CJ, Liu HW (agosto de 2007). "Los diversos roles de las coenzimas flavinas - los actores más versátiles de la naturaleza" . La Revista de Química Orgánica . 72 (17): 6329–42. doi : 10.1021 / jo0703092 . PMC 2519020 . PMID 17580897 .
- ^ Rey MW. "Vitaminas, Minerales, Suplementos" . La página de bioquímica médica .
- ↑ Garma, Leonardo D .; Medina, Milagros; Juffer, André H. (1 de noviembre de 2016). "Clasificación basada en la estructura de los sitios de unión de FAD: un estudio comparativo de herramientas de alineación estructural". Proteínas: estructura, función y bioinformática . 84 (11): 1728-1747. doi : 10.1002 / prot.25158 . ISSN 1097-0134 . PMID 27580869 . S2CID 26066208 .
- ↑ a b c d e f g h Fagan RL, Palfey BA (2010). "Enzimas dependientes de flavina". Productos Naturales Integrales II Química y Biología . 7 : 37-113.
- ↑ a b Hanukoglu I (1996). "Proteínas de transferencia de electrones de los sistemas del citocromo P450" (PDF) . Adv. Mol. Cell Biol . Avances en Biología Molecular y Celular. 14 : 29–55. doi : 10.1016 / S1569-2558 (08) 60339-2 . ISBN 9780762301133.
- ^ Ziegler GA, Vonrhein C, Hanukoglu I, Schulz GE (junio de 1999). "La estructura de la adrenodoxina reductasa de los sistemas mitocondriales P450: transferencia de electrones para la biosíntesis de esteroides". Revista de Biología Molecular . 289 (4): 981–90. doi : 10.1006 / jmbi.1999.2807 . PMID 10369776 .
- ↑ a b Hanukoglu I (2017). "Conservación de las interfaces enzima-coenzima en FAD y NADP que se une a la enzima ubicua de adrenodoxina reductasa-A". Revista de evolución molecular . 85 (5): 205–218. doi : 10.1007 / s00239-017-9821-9 . PMID 29177972 . S2CID 7120148 .
- ^ Hanukoglu I, Jefcoate CR (abril de 1980). "Citocromo P-450scc mitocondrial. Mecanismo de transporte de electrones por adrenodoxina" (PDF) . La revista de química biológica . 255 (7): 3057–61. doi : 10.1016 / S0021-9258 (19) 85851-9 . PMID 6766943 .
- ^ McNeil MB, Fineran PC (mayo de 2013). "Factores de ensamblaje procariotas para la unión de flavina al complejo II" . Biochimica et Biophysica Acta (BBA) - Bioenergética . 1827 (5): 637–47. doi : 10.1016 / j.bbabio.2012.09.003 . PMID 22985599 .
- ^ Serrano A, Ferreira P, Martínez-Júlvez M, Medina M (2013). "La familia procariota FAD sintetasa: un potencial objetivo de la droga". Diseño Farmacéutico Actual . 19 (14): 2637–48. doi : 10.2174 / 1381612811319140013 . PMID 23116401 .
- ^ a b c d Christie JM, Gawthorne J, Young G, Fraser NJ, Roe AJ (mayo de 2012). "LOV a BLUF: contribuciones de flavoproteínas al conjunto de herramientas optogenéticas" . Planta molecular . 5 (3): 533–44. doi : 10.1093 / mp / sss020 . PMID 22431563 .
- ↑ a b c Sivabalan S, Vedeswari CP, Jayachandran S, Koteeswaran D, Pravda C, Aruna PR, Ganesan S (2010). "Espectroscopia de fluorescencia nativa in vivo y reducción de dinucleótido de nicotinamida adinina / dinucleótido de flavina adenina y estados de oxidación de la fibrosis submucosa oral para la monitorización de fármacos quimiopreventivos". Revista de Óptica Biomédica . 15 (1): 017010. doi : 10.1117 / 1.3324771 . PMID 20210484 .
Enlaces externos
- FAD unido a proteínas en el AP
- Entrada FAD en la base de datos química de los NIH