Na v 1.8 es un subtipo de canal de iones de sodio que en humanos está codificado por el gen SCN10A . [5] [6] [7] [8]
Los canales que contienen Na v 1.8 son canales dependientes de voltaje resistentes a tetrodotoxina (TTX). Na v 1.8 se expresa específicamente en el ganglio de la raíz dorsal (DRG), en neuronas sensoriales amielínicas de pequeño diámetro llamadas fibras C , y participa en la nocicepción . [9] [10] Las fibras C pueden ser activadas por estímulos térmicos o mecánicos nocivos y, por lo tanto, pueden transmitir mensajes de dolor .
La ubicación específica de Na v 1.8 en las neuronas sensoriales de los GRD puede convertirlo en un objetivo terapéutico clave para el desarrollo de nuevos analgésicos [11] y el tratamiento del dolor crónico . [12]
Función
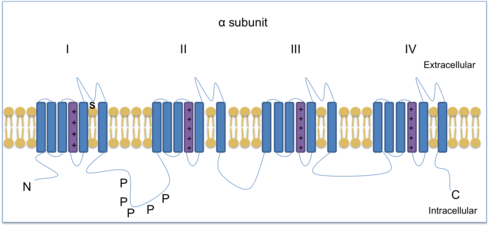
Los canales de iones de sodio activados por voltaje (VGSC) son esenciales para producir y propagar potenciales de acción . La tetrodotoxina, una toxina que se encuentra en el pez globo , puede bloquear algunos VGSC y, por lo tanto, se utiliza para distinguir los diferentes subtipos. Hay tres VGSC resistentes a TTX: Na v 1.5 , Na v 1.8 y Na v 1.9 . Na v 1.8 y Na v 1.9 se expresan ambos en nociceptores (neuronas sensibles al daño). Na v 1.7 , Na v 1.8 y Na v 1.9 se encuentran en el DRG y ayudan a mediar el dolor inflamatorio crónico.[13] Na v 1.8 es una subunidad de canal de tipo α que consta de cuatro dominios homólogos, cada uno con seis regiones transmembrana, de las cuales una es un sensor de voltaje.
Estructura de Na v 1.8, una subunidad de tipo α con cuatro dominios homólogos, cada uno con seis regiones transmembrana. Cada dominio tiene un sensor de voltaje (violeta). La 'P' representa los sitios de fosforilación de la proteína quinasa A ; N y C indican los extremos amino y carboxi de la cadena proteica. Esta imagen ha sido adaptada de 'El tráfico de Na v 1.8' [12]
Los métodos de fijación de voltaje han demostrado que el Na V 1.8 es único, entre los canales de sodio, al exhibir una inactivación en estado estacionario relativamente despolarizada. Así, el Na V 1.8 permanece disponible para operar, cuando las neuronas se despolarizan a niveles que inactivan otros canales de sodio. La pinza de voltaje se ha utilizado para mostrar cómo los potenciales de acción en las células DRG se moldean mediante canales de sodio resistentes a TTX. Na v 1.8 es el que más contribuye a mantener la etapa despolarizante de los potenciales repetitivos de alta frecuencia de acción en las neuronas sensoriales nociceptivas porque se activa rápidamente y permanece activa después de detectar un estímulo nocivo . [14] [15] Por lo tanto, Na v 1.8 contribuye ahiperalgesia (aumento de la sensibilidad al dolor) y alodinia (dolor por estímulos que no suelen causarlo), que son elementos del dolor crónico. [16] Estudios en ratones knockout Na v 1.8 han demostrado que el canal está asociado con dolor inflamatorio y neuropático. [9] [17] [18] Además, Na v 1.8 juega un papel crucial en el dolor por frío. [19] Reducir la temperatura de 30 ° C a 10 ° C ralentiza la activación de los VGSC y, por lo tanto, disminuye la corriente. Sin embargo, Na v 1.8 es resistente al frío y es capaz de generar potenciales de acción en el frío para llevar información de los nociceptores alsistema nervioso central (SNC). Además, los ratones nulos para Na v 1.8 no produjeron potenciales de acción, lo que indica que Na v 1.8 es esencial para la percepción del dolor en temperaturas frías. [19]
Aunque los primeros estudios sobre la biofísica de los canales de Na V 1.8 se llevaron a cabo en canales de roedores, estudios más recientes han examinado las propiedades de los canales de Na V 1.8 humanos . En particular, los canales de Na V 1.8 humanos exhiben una dependencia del voltaje de inactivación que está incluso más despolarizada que la de los roedores, y también exhibe una corriente persistente más grande. [20] Por lo tanto, la influencia de los canales de Na V 1.8 humanos en la activación de las neuronas sensoriales puede ser incluso mayor que la de los canales de Na V 1.8 de los roedores .
Se ha descubierto que las mutaciones de ganancia de función de Na V 1.8, identificadas en pacientes con neuropatías periféricas dolorosas, hacen que las neuronas DRG sean hiperexcitables y, por lo tanto, son causas de dolor. [21] [22] Aunque Na V 1.8 normalmente no se expresa dentro del cerebelo, su expresión está regulada por incremento en las células cerebelosas de Purkinje en modelos animales de EM (esclerosis múltiple) y en seres humanos. [23] La presencia de canales Na V 1.8 dentro de estas neuronas cerebelosas, donde normalmente no está presente, aumenta su excitabilidad y altera su patrón de activación in vitro, [24] y en roedores con encefalomielitis autoinmune experimental, un modelo de EM. [25]A nivel conductual, se ha demostrado que la expresión ectópica de Na V 1.8 dentro de las neuronas cerebelosas de Purkinje perjudica el rendimiento motor en un modelo transgénico. [26]
Importancia clínica
Vías de señalización del dolor
Los nociceptores se diferencian de otras neuronas sensoriales en que tienen un umbral de activación bajo y, en consecuencia, aumentan su respuesta a estímulos constantes. Por lo tanto, los nociceptores se sensibilizan fácilmente por agentes como la bradicinina y el factor de crecimiento nervioso , que se liberan en el sitio de la lesión tisular y, en última instancia, provocan cambios en la conductancia del canal iónico. Se ha demostrado que las VGSC aumentan de densidad después de una lesión nerviosa. [27] Por lo tanto, las VGSC pueden ser moduladas por muchos agentes hiperalgésicos diferentes que se liberan después de una lesión nerviosa. Otros ejemplos incluyen prostaglandina E 2 (PGE 2 ), serotonina y adenosina, que actúan todos para aumentar la corriente a través de Na v 1.8. [28]
Las prostaglandinas como la PGE 2 pueden sensibilizar a los nociceptores a estímulos térmicos, químicos y mecánicos y aumentar la excitabilidad de las neuronas sensoriales DRG. Esto ocurre porque PGE 2 modula el tráfico de Na v 1,8 mediante la unión a acoplado a la proteína G del receptor EP2 , que a su vez activa la proteína quinasa A . [29] [30] La proteína quinasa A fosforila el Na v 1.8 en los sitios intracelulares, lo que aumenta las corrientes de iones de sodio. La evidencia de un vínculo entre la PGE 2 y la hiperalgesia proviene de una eliminación de desoxinucleótido antisentido de Na v 1.8 en el DRG de ratas. [31] Otro modulador de Nav 1.8 es la isoforma ε de PKC . Esta isoforma es activada por el mediador inflamatorio bradicinina y fosforila Na v 1.8, provocando un aumento de la corriente de sodio en las neuronas sensoriales, lo que favorece la hiperalgesia mecánica. [32]
Síndrome de Brugada
Las mutaciones en SCN10A están asociadas con el síndrome de Brugada . [33] [34] [35]
Tráfico de membranas
Los niveles del factor de crecimiento nervioso en los tejidos inflamados o lesionados aumentan creando una mayor sensibilidad al dolor (hiperalgesia). [36] El aumento de los niveles de factor de crecimiento nervioso y factor de necrosis tumoral α (TNF-α) provoca la regulación positiva de Na v 1.8 en las neuronas sensoriales a través de la proteína accesoria p11 (cadena ligera de la anexina II). Se ha demostrado usando el método de selección de híbridos de levadura dos que p11 se une a un fragmento de 28 aminoácidos en el extremo N de Na v 1.8 y promueve su translocación a la membrana plasmática . Esto contribuye a la hiperexcitabilidad de las neuronas sensoriales durante el dolor. [37]neuronas p11 nulo nociceptivas sensoriales en ratones, creado usando el Cre- loxP recombinasa sistema, muestran una disminución en Na v 1,8 expresión en la membrana plasmática. [38] Por lo tanto, interrumpir las interacciones entre p11 y Na v 1.8 puede ser un buen objetivo terapéutico para reducir el dolor.
En las fibras mielinizadas , las VGSC se encuentran en los nodos de Ranvier ; sin embargo, en fibras amielínicas, no se ha determinado la ubicación exacta de las VGSC. Se ha encontrado Na v 1.8 en fibras amielínicas en grupos asociados con balsas lipídicas a lo largo de las fibras DRG tanto in vitro como in vivo . [39] Las balsas de lípidos organizan la membrana celular, que incluye el tráfico y la localización de canales iónicos. La eliminación de balsas de lípidos en la membrana utilizando MβCD , que agota el colesterol de la membrana plasmática, conduce a un cambio de Na v1.8 a una parte de la membrana que no es balsa, lo que reduce el potencial de acción de disparo y propagación. [39]
Neuropatías periféricas dolorosas
Las neuropatías periféricas dolorosas o las neuropatías de fibras pequeñas son trastornos de las fibras C nociceptivas amielínicas que causan dolor neuropático; en algunos casos, no hay una causa conocida. [40] El examen genético de pacientes con estas neuropatías idiopáticas ha descubierto mutaciones en el gen SCN9A , que codifica el canal relacionado Na v 1.7. Se encontró una mutación de ganancia de función en Na v 1.7 ubicada en las neuronas sensoriales del DRG en el 30% de los pacientes. [41] Esta mutación de ganancia de función provoca un aumento de la excitabilidad (hiperexcitabilidad) de las neuronas sensoriales de los DRG y, por lo tanto, un aumento del dolor. Na v1.7 así se ha demostrado que está relacionado con el dolor humano; Na v 1.8, por el contrario, solo se había asociado con el dolor en estudios con animales hasta hace poco. Se encontró una mutación de ganancia de función en el gen SCN10A que codifica Na v 1.8 en pacientes con neuropatía periférica dolorosa. [21] En un estudio de 104 pacientes con neuropatías periféricas idiopáticas que no tenían la mutación en SCN9A, se utilizaron métodos de fijación de voltaje y fijación de corriente , junto con algoritmos predictivos , y arrojó dos mutaciones de ganancia de función en SCN10Aen tres pacientes. Ambas mutaciones causan un aumento de la excitabilidad en las neuronas sensoriales de los DRG y, por lo tanto, contribuyen al dolor, pero no se comprende el mecanismo por el cual lo hacen.
Referencias
^ a b c GRCh38: Ensembl release 89: ENSG00000185313 - Ensembl , mayo de 2017
^ a b c GRCm38: Ensembl release 89: ENSMUSG00000034533 - Ensembl , mayo de 2017
^ "Referencia humana de PubMed:" . Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ "Referencia de PubMed del ratón:" . Centro Nacional de Información Biotecnológica, Biblioteca Nacional de Medicina de EE. UU .
^ "Entrez Gene: canal de sodio" .
^ Rabert DK, Koch BD, Ilnicka M, Obernolte RA, Naylor SL, Herman RC, Eglen RM, Hunter JC, Sangameswaran L (noviembre de 1998). "Un canal de sodio dependiente de voltaje resistente a la tetrodotoxina de los ganglios de la raíz dorsal humana, hPN3 / SCN10A". Dolor . 78 (2): 107-14. doi : 10.1016 / S0304-3959 (98) 00120-1 . PMID 9839820 . S2CID 45480324 .
^ Plummer NW, Meisler MH (abril de 1999). "Evolución y diversidad de genes de canales de sodio de mamíferos". Genómica . 57 (2): 323–31. doi : 10.1006 / geno.1998.5735 . PMID 10198179 .
^ Catterall WA, Goldin AL, Waxman SG (diciembre de 2005). "Unión Internacional de Farmacología. XLVII. Nomenclatura y relaciones estructura-función de los canales de sodio dependientes de voltaje". Revisiones farmacológicas . 57 (4): 397–409. doi : 10.1124 / pr.57.4.4 . PMID 16382098 . S2CID 7332624 .
^ a b Akopian AN, Souslova V, England S, Okuse K, Ogata N, Ure J, Smith A, Kerr BJ, McMahon SB, Boyce S, Hill R, Stanfa LC, Dickenson AH, Wood JN (junio de 1999). "El SNS del canal de sodio resistente a la tetrodotoxina tiene una función especializada en las vías del dolor". Neurociencia de la naturaleza . 2 (6): 541–8. doi : 10.1038 / 9195 . PMID 10448219 . S2CID 17487906 .
^ Akopian AN, Sivilotti L, Wood JN (enero de 1996). "Un canal de sodio dependiente de voltaje resistente a la tetrodotoxina expresado por neuronas sensoriales". Naturaleza . 379 (6562): 257–62. Código Bibliográfico : 1996Natur.379..257A . doi : 10.1038 / 379257a0 . PMID 8538791 . S2CID 4360775 .
^ Cummins TR, Sheets PL, Waxman SG (octubre de 2007). "Los roles de los canales de sodio en la nocicepción: implicaciones para los mecanismos del dolor" . Dolor . 131 (3): 243–57. doi : 10.1016 / j.pain.2007.07.026 . PMC 2055547 . PMID 17766042 .
↑ a b Swanwick RS, Pristerá A, Okuse K (diciembre de 2010). "El tráfico de Na (V) 1.8" . Cartas de neurociencia . 486 (2): 78–83. doi : 10.1016 / j.neulet.2010.08.074 . PMC 2977848 . PMID 20816723 .
^ Strickland IT, Martindale JC, Woodhams PL, Reeve AJ, Chessell IP, McQueen DS (julio de 2008). "Cambios en la expresión de NaV1.7, NaV1.8 y NaV1.9 en una población distinta de ganglios de la raíz dorsal que inervan la articulación de la rodilla de rata en un modelo de dolor articular inflamatorio crónico". Revista europea del dolor . 12 (5): 564–72. doi : 10.1016 / j.ejpain.2007.09.001 . PMID 17950013 . S2CID 24952010 .
^ Blair NT, Bean BP (2002). "Funciones de la corriente de Na + sensible a la tetrodotoxina (TTX), la corriente de Na + resistente a TTX y la corriente de Ca2 + en los potenciales de acción de las neuronas sensoriales nociceptivas" . La Revista de Neurociencia . 22 (23): 10277–10290. doi : 10.1523 / JNEUROSCI.22-23-10277.2002 . PMC 6758735 . PMID 12451128 .
^ Renganathan M, Cummins TR y Waxman SG (2001). "Contribución de los canales de sodio Nav1.8 a la electrogénesis potencial de acción en las neuronas DRG". Revista de neurofisiología . 86 (2): 629–640. doi : 10.1152 / jn.2001.86.2.629 . PMID 11495938 .
^ Millan MJ (1999). "La inducción del dolor: una revisión integradora". Avances en neurobiología . 57 (1): 1-164. doi : 10.1016 / S0301-0082 (98) 00048-3 . PMID 9987804 . S2CID 206054345 .
^ Matthews EA, Wood JN, Dickenson AH (febrero de 2006). "Los ratones nulos Na (v) 1.8 muestran déficits dependientes de estímulos en la actividad neuronal espinal" . Dolor molecular . 2 : 1744-8069–2-5. doi : 10.1186 / 1744-8069-2-5 . PMC 1403745 . PMID 16478543 .
^ Jarvis MF, Honore P, Shieh CC, Chapman M, Joshi S, Zhang XF, Kort M, Carroll W, Marron B, Atkinson R, Thomas J, Liu D, Krambis M, Liu Y, McGaraughty S, Chu K, Roeloffs R, Zhong C, Mikusa JP, Hernandez G, Gauvin D, Wade C, Zhu C, Pai M, Scanio M, Shi L, Drizin I, Gregg R, Matulenko M, Hakeem A, Gross M, Johnson M, Marsh K, Wagoner PK, Sullivan JP, Faltynek CR, Krafte DS (mayo de 2007). "A-803467, un bloqueador de los canales de sodio Nav1.8 potente y selectivo, atenúa el dolor neuropático e inflamatorio en la rata" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 104 (20): 8520–5. doi : 10.1073 / pnas.0611364104 . PMC 1895982 . PMID 17483457 .
↑ a b Zimmermann K, Leffler A, Babes A, Cendan CM, Carr RW, Kobayashi J, Nau C, Wood JN, Reeh PW (junio de 2007). "El canal de sodio de la neurona sensorial Nav1.8 es esencial para el dolor a bajas temperaturas". Naturaleza . 447 (7146): 855–8. Código Bibliográfico : 2007Natur.447..856Z . doi : 10.1038 / nature05880 . PMID 17568746 . S2CID 4391511 .
^ Han C, Estación M, Huang J, Vasylyev D, Zhao P, Dib-Hajj SD, Waxman SG (mayo de 2015). "Human Na (v) 1.8: las corrientes persistentes y en rampa mejoradas contribuyen a distintas propiedades de disparo de las neuronas DRG humanas" . Revista de neurofisiología . 113 (9): 3172–85. doi : 10.1152 / jn.00113.2015 . PMC 4432682 . PMID 25787950 .
^ a b Faber CG, Lauria G, Merkies IS, Cheng X, Han C, Ahn HS, Persson AK, Hoeijmakers JG, Gerrits MM, Pierro T, Lombardi R, Kapetis D, Dib-Hajj SD, Waxman SG (noviembre de 2012) . "Mutaciones de ganancia de función Nav1.8 en neuropatía dolorosa" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 109 (47): 19444–9. Código bibliográfico : 2012PNAS..10919444F . doi : 10.1073 / pnas.1216080109 . PMC 3511073 . PMID 23115331 .
^ Huang J, Yang Y, Zhao P, Gerrits MM, Hoeijmakers JG, Bekelaar K, Merkies IS, Faber CG, Dib-Hajj SD, Waxman SG (agosto de 2013). "La mutación Nav1.8 de neuropatía de fibras pequeñas cambia la activación a potenciales hiperpolarizados y aumenta la excitabilidad de las neuronas del ganglio de la raíz dorsal" . La Revista de Neurociencia . 33 (35): 14087–97. doi : 10.1523 / JNEUROSCI.2710-13.2013 . PMC 6618513 . PMID 23986244 .
^ Black JA, Dib-Hajj S, Baker D, Newcombe J, Cuzner ML, Waxman SG (octubre de 2000). "SNS del canal de sodio específico de neuronas sensoriales se expresa anormalmente en los cerebros de ratones con encefalomielitis alérgica experimental y humanos con esclerosis múltiple" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 97 (21): 11598–602. Código bibliográfico : 2000PNAS ... 9711598B . doi : 10.1073 / pnas.97.21.11598 . PMC 17246 . PMID 11027357 .
^ Renganathan M, Gelderblom M, Black JA, Waxman SG (enero de 2003). "La expresión de los canales de sodio Nav1.8 perturba los patrones de activación de las células cerebelosas de Purkinje". Investigación del cerebro . 959 (2): 235–42. doi : 10.1016 / s0006-8993 (02) 03750-2 . PMID 12493611 . S2CID 34784900 .
^ Saab CY, Craner MJ, Kataoka Y, Waxman SG (septiembre de 2004). "Actividad anormal de las células de Purkinje in vivo en encefalomielitis alérgica experimental". Investigación experimental del cerebro . 158 (1): 1–8. doi : 10.1007 / s00221-004-1867-4 . PMID 15118796 . S2CID 34656521 .
^ Shields SD, Cheng X, Gasser A, Saab CY, Tyrrell L, Eastman EM, Iwata M, Zwinger PJ, Black JA, Dib-Hajj SD, Waxman SG (febrero de 2012). "Una canalopatía contribuye a la disfunción cerebelosa en un modelo de esclerosis múltiple". Annals of Neurology . 71 (2): 186–94. doi : 10.1002 / ana.22665 . PMID 22367990 . S2CID 25128887 .
^ Devor M; Govrin-Lippmann R y Angelides (1993). "Inmunolocalización del canal de Na + en axones de mamíferos periféricos y cambios tras la formación de lesiones nerviosas y neuromas" . La Revista de Neurociencia . 13 (5): 1976–1992. doi : 10.1523 / JNEUROSCI.13-05-01976.1993 . PMC 6576562 . PMID 7683047 .
^ Gold MS, Reichling DB, Shuster MJ, Levine JD (febrero de 1996). "Los agentes hiperalgésicos aumentan una corriente de Na + resistente a tetrodotoxina en los nociceptores" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 93 (3): 1108–12. Código Bibliográfico : 1996PNAS ... 93.1108G . doi : 10.1073 / pnas.93.3.1108 . PMC 40039 . PMID 8577723 .
^ Héctor TH (enero de 1975). "Un método sencillo para realizar registros cromatográficos utilizando una hoja de acetato transparente" . La revista de fisiología . 32 (1): 31–2. doi : 10.1113 / jphysiol.1996.sp021604 . PMC 1160802 . PMID 8887754 .
^ Liu C, Li Q, Su Y, Bao L (marzo de 2010). "La prostaglandina E2 promueve el tráfico de Na1.8 a través de su motivo RRR intracelular a través de la ruta de la proteína quinasa A" . Tráfico . 11 (3): 405-17. doi : 10.1111 / j.1600-0854.2009.01027.x . PMID 20028484 . S2CID 997800 .
^ Khasar SG, Gold MS y Levine JD (1998). "Una corriente de sodio resistente a la tetrodotoxina media el dolor inflamatorio en la rata" . Cartas de neurociencia . 256 (1): 17-20. doi : 10.1016 / s0304-3940 (98) 00738-1 . PMID 9832206 . S2CID 5614913 .
^ Wu DF, Chandra D, McMahon T, Wang D, Dadgar J, Kharazia VN, Liang YJ, Waxman SG, Dib-Hajj SD, Messing RO (abril de 2012). "La fosforilación de PKCε del canal de sodio NaV1.8 aumenta la función del canal y produce hiperalgesia mecánica en ratones" . La Revista de Investigación Clínica . 122 (4): 1306–15. doi : 10.1172 / JCI61934 . PMC 3315445 . PMID 22426212 .
^ Hu D, Barajas-Martínez H, Pfeiffer R, Dezi F, Pfeiffer J, Buch T, Betzenhauser MJ, Belardinelli L, Kahlig KM, Rajamani S, DeAntonio HJ, Myerburg RJ, Ito H, Deshmukh P, Marieb M, Nam GB , Bhatia A, Hasdemir C, Haïssaguerre M, Veltmann C, Schimpf R, Borggrefe M, Viskin S, Antzelevitch C (julio de 2014). "Las mutaciones en SCN10A son responsables de una gran fracción de los casos de síndrome de Brugada" . Revista del Colegio Americano de Cardiología . 64 (1): 66–79. doi : 10.1016 / j.jacc.2014.04.032 . PMC 4116276 . PMID 24998131 .
^ Monasky MM, Micaglio E, Vicedomini G, Locati ET, Ciconte G, Giannelli L, Giordano F, Crisà S, Vecchi M, Borrelli V, Ghiroldi A, D'Imperio S, Di Resta C, Benedetti S, Ferrari M, Santinelli V, Anastasia L, Pappone C (2019). "Características clínicas comparables en pacientes con síndrome de Brugada que albergan SCN5A o variantes nuevas de SCN10A" . Europace . 21 (10): 1550-1558. doi : 10.1093 / europace / euz186 . PMID 31292628 . Consultado el 27 de abril de 2021 .CS1 maint: multiple names: authors list (link)
^ Monasky MM, Micaglio E, Ciconte G, Pappone C (2020). "Síndrome de Brugada: ¿enfermedad oligogénica o mendeliana?" . Int J Mol Sci . 21 (5): 1687. doi : 10.3390 / ijms21051687 . PMID 32121523 . Consultado el 27 de abril de 2021 .CS1 maint: multiple names: authors list (link)
^ McMahon SB (marzo de 1996). "NGF como mediador del dolor inflamatorio". Transacciones filosóficas de la Royal Society de Londres. Serie B, Ciencias Biológicas . 351 (1338): 431–40. Código Bibliográfico : 1996RSPTB.351..431M . doi : 10.1098 / rstb.1996.0039 . PMID 8730782 .
^ Okuse K, Malik-Hall M, Baker MD, Poon WY, Kong H, Chao MV, Wood JN (junio de 2002). "La cadena ligera de anexina II regula la expresión del canal de sodio específico de neuronas sensoriales". Naturaleza . 417 (6889): 653–6. Código Bibliográfico : 2002Natur.417..653O . doi : 10.1038 / nature00781 . PMID 12050667 . S2CID 4423351 .
^ Foulkes T, Nassar MA, Lane T, Matthews EA, Baker MD, Gerke V, Okuse K, Dickenson AH, Wood JN (octubre de 2006). "La eliminación de la cadena ligera de la anexina 2 p11 en los nociceptores provoca déficits en la codificación somatosensorial y el comportamiento del dolor" (PDF) . La Revista de Neurociencia . 26 (41): 10499–507. doi : 10.1523 / JNEUROSCI.1997-06.2006 . PMC 6674704 . PMID 17035534 .
↑ a b Pristerà A, Baker MD, Okuse K (2012). "La asociación entre los canales resistentes a la tetrodotoxina y las balsas lipídicas regula la excitabilidad de las neuronas sensoriales" . PLOS ONE . 7 (8): e40079. Código bibliográfico : 2012PLoSO ... 740079P . doi : 10.1371 / journal.pone.0040079 . PMC 3411591 . PMID 22870192 .
^ Hoeijmakers JG, Faber CG, Lauria G, Merkies IS, Waxman SG (mayo de 2012). "Neuropatías de fibras pequeñas: avances en diagnóstico, fisiopatología y tratamiento". Reseñas de la naturaleza. Neurología . 8 (7): 369–79. doi : 10.1038 / nrneurol.2012.97 . PMID 22641108 . S2CID 8804151 .
^ Faber CG, Hoeijmakers JG, Ahn HS, Cheng X, Han C, Choi JS, Estacion M, Lauria G, Vanhoutte EK, Gerrits MM, Dib-Hajj S, Drenth JP, Waxman SG, Merkies IS (enero de 2012). "Ganancia de mutaciones de función Naν1.7 en neuropatía idiopática de fibras pequeñas". Annals of Neurology . 71 (1): 26–39. doi : 10.1002 / ana.22485 . PMID 21698661 . S2CID 11711575 .
Lectura adicional
Okuse K (2007). "Vías de señalización del dolor: de citocinas a canales iónicos". La Revista Internacional de Bioquímica y Biología Celular . 39 (3): 490–6. doi : 10.1016 / j.biocel.2006.11.016 . PMID 17194618 .
Waxman SG (julio de 2013). "Canalopatías de Na dolorosas: un universo en expansión". Tendencias en Medicina Molecular . 19 (7): 406–9. doi : 10.1016 / j.molmed.2013.04.003 . PMID 23664154 .
Lai J, Porreca F, Hunter JC, Gold MS (2004). "Canales de sodio dependientes de voltaje e hiperalgesia". Revista anual de farmacología y toxicología . 44 : 371–97. doi : 10.1146 / annurev.pharmtox.44.101802.121627 . PMID 14744251 .
Wood JN, Boorman JP, Okuse K, Baker MD (octubre de 2004). "Canales de sodio dependientes de voltaje y vías del dolor". Revista de neurobiología . 61 (1): 55–71. doi : 10.1002 / neu.20094 . PMID 15362153 .
Malik-Hall M, Poon WY, Baker MD, Wood JN, Okuse K (febrero de 2003). "Las proteínas de las neuronas sensoriales interactúan con los dominios intracelulares del canal de sodio NaV1.8". Investigación del cerebro. Investigación del cerebro molecular . 110 (2): 298-304. doi : 10.1016 / S0169-328X (02) 00661-7 . PMID 12591166 .
Yamaoka K, Inoue M, Miyazaki K, Hirama M, Kondo C, Kinoshita E, Miyoshi H, Seyama I (marzo de 2009). "Las ciguatoxinas sintéticas activan selectivamente los canales de sodio quiméricos derivados de Nav1.8 expresados en células HEK293" . La revista de química biológica . 284 (12): 7597–605. doi : 10.1074 / jbc.M806481200 . PMC 2658054 . PMID 19164297 .
Choi JS, Hudmon A, Waxman SG, Dib-Hajj SD (julio de 2006). "Calmodulin regula la densidad de corriente y la inhibición dependiente de la frecuencia del canal de sodio Nav1.8 en neuronas DRG". Revista de neurofisiología . 96 (1): 97–108. doi : 10.1152 / jn.00854.2005 . PMID 16598065 .
Liu CJ, Priest BT, Bugianesi RM, Dulski PM, Felix JP, Dick IE, Brochu RM, Knaus HG, Middleton RE, Kaczorowski GJ, Slaughter RS, Garcia ML, Köhler MG (febrero de 2006). "Un ensayo basado en FRET de potencial de membrana de alta capacidad para canales NaV1.8". Tecnologías de análisis y desarrollo de fármacos . 4 (1): 37–48. doi : 10.1089 / adt.2006.4.37 . PMID 16506887 .
Browne LE, Blaney FE, Yusaf SP, Clare JJ, Wray D (abril de 2009). "Determinantes estructurales de las drogas que actúan sobre el canal Nav1.8" . La revista de química biológica . 284 (16): 10523–36. doi : 10.1074 / jbc.M807569200 . PMC 2667739 . PMID 19233853 .
Rabert DK, Koch BD, Ilnicka M, Obernolte RA, Naylor SL, Herman RC, Eglen RM, Hunter JC, Sangameswaran L (noviembre de 1998). "Un canal de sodio dependiente de voltaje resistente a la tetrodotoxina de los ganglios de la raíz dorsal humana, hPN3 / SCN10A". Dolor . 78 (2): 107-14. doi : 10.1016 / S0304-3959 (98) 00120-1 . PMID 9839820 . S2CID 45480324 .
Plummer NW, Meisler MH (abril de 1999). "Evolución y diversidad de genes de canales de sodio de mamíferos". Genómica . 57 (2): 323–31. doi : 10.1006 / geno.1998.5735 . PMID 10198179 .
Catterall WA, Goldin AL, Waxman SG (diciembre de 2005). "Unión Internacional de Farmacología. XLVII. Nomenclatura y relaciones estructura-función de los canales de sodio dependientes de voltaje". Revisiones farmacológicas . 57 (4): 397–409. doi : 10.1124 / pr.57.4.4 . PMID 16382098 . S2CID 7332624 .
Enlaces externos
El Consorcio del Dolor de Londres
vtmiProteína de transporte de membrana : canales iónicos ( TC 1A )