El color de las hojas otoñales es un fenómeno que afecta a las hojas verdes normales de muchos árboles y arbustos caducifolios , por lo que adquieren, durante algunas semanas de la temporada otoñal, diversas tonalidades de amarillo, naranja, rojo, violeta y marrón. [1] El fenómeno se denomina comúnmente colores otoñales [2] o follaje otoñal [3] en inglés británico y colores otoñales , [4] follaje otoñal , o simplemente follaje [5] en inglés estadounidense .



En algunas áreas de Canadá y los Estados Unidos , el turismo de " observación de hojas " es una contribución importante a la actividad económica. Esta actividad turística ocurre entre el inicio de los cambios de color y el inicio de la caída de las hojas , generalmente alrededor de septiembre y octubre en el hemisferio norte y de abril a mayo en el hemisferio sur .
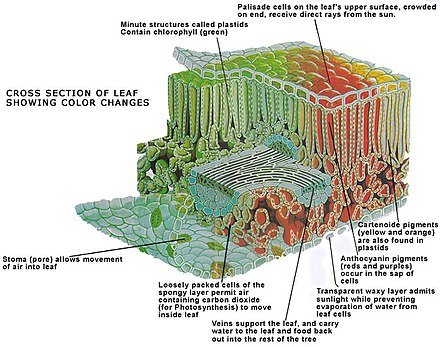
Clorofila y los colores verde / amarillo / naranja
.jpg/440px-Autumn_colours_in_Heinola,_Finland_(October_2018).jpg)
Una hoja verde es verde debido a la presencia de un pigmento conocido como clorofila , que se encuentra dentro de un orgánulo llamado cloroplasto . Cuando abunda en las células de la hoja , como durante la temporada de crecimiento, el color verde de la clorofila domina y enmascara los colores de cualquier otro pigmento que pueda estar presente en la hoja. Así, las hojas del verano son característicamente verdes. [6]
La clorofila tiene una función vital: captura los rayos solares y utiliza la energía resultante en la fabricación de los alimentos de la planta: azúcares simples que se producen a partir del agua y el dióxido de carbono . Estos azúcares son la base de la nutrición de la planta, la única fuente de carbohidratos necesarios para el crecimiento y el desarrollo. En su proceso de fabricación de alimentos, las clorofilas se descomponen, por lo que se "agotan" continuamente. Sin embargo, durante la temporada de crecimiento, la planta repone la clorofila para que el suministro se mantenga alto y las hojas se mantengan verdes.
A medida que se acerca el otoño, con el acortamiento de las horas de luz y el enfriamiento de las temperaturas, las venas que llevan los fluidos hacia y desde la hoja se cierran gradualmente a medida que se forma una capa de células especiales de corcho en la base de cada hoja. A medida que se desarrolla esta capa de corcho, la ingesta de agua y minerales en la hoja se reduce, primero lentamente y luego más rápidamente. Durante este tiempo, la cantidad de clorofila en la hoja comienza a disminuir. A menudo, las venas todavía están verdes después de que los tejidos entre ellas hayan cambiado casi por completo de color.
Mucha clorofila se encuentra en el fotosistema II (complejo de captación de luz II o LHC II), la proteína de membrana más abundante en la tierra. [ cita requerida ] LHC II captura la luz en la fotosíntesis. Se encuentra en la membrana tilacoide del cloroplasto y está compuesto por una apoproteína junto con varios ligandos , siendo los más importantes las clorofilas ay b. En otoño, este complejo se descompone. Se cree que la degradación de la clorofila ocurre primero. Las investigaciones sugieren que el comienzo de la degradación de la clorofila es catalizado por la clorofila b reductasa, que reduce la clorofila b a 7-hidroximetil clorofila a, que luego se reduce a clorofila a. [7]Se cree que esto desestabiliza el complejo, momento en el que se produce la degradación de la apoproteína. Una enzima importante en la degradación de la apoproteína es FtsH6, que pertenece a la familia de proteasas FtsH . [8]
Las clorofilas se degradan en tetrapirroles incoloros conocidos como catabolitos de clorofila no fluorescentes. [9] A medida que se degradan las clorofilas, se revelan los pigmentos ocultos de las xantofilas amarillas y el betacaroteno naranja . Estos pigmentos están presentes durante todo el año, pero los pigmentos rojos, las antocianinas , se sintetizan de novo una vez que se ha degradado aproximadamente la mitad de la clorofila. Los aminoácidos liberados por la degradación de los complejos de recolección de luz se almacenan durante todo el invierno en las raíces, ramas, tallos y tronco del árbol hasta la próxima primavera, cuando se reciclan para liberar el árbol. [ cita requerida]
Pigmentos que contribuyen a otros colores
Carotenoides
Los carotenoides están presentes en las hojas durante todo el año, pero sus colores amarillo anaranjado suelen estar enmascarados por la clorofila verde. [6] A medida que se acerca el otoño, ciertas influencias tanto dentro como fuera de la planta hacen que las clorofilas sean reemplazadas a un ritmo más lento de lo que se están agotando. Durante este período, con el suministro total de clorofilas disminuyendo gradualmente, el efecto de "enmascaramiento" se desvanece lentamente. Luego, otros pigmentos presentes (junto con las clorofilas) en las células de la hoja comienzan a verse. [6] Estos son carotenoides y proporcionan coloraciones de amarillo, marrón, naranja y los muchos matices intermedios.
Los carotenoides se encuentran, junto con los pigmentos de clorofila, en estructuras diminutas llamadas plastidios , dentro de las células de las hojas. A veces, son tan abundantes en la hoja que le dan a la planta un color amarillo verdoso, incluso durante el verano. Por lo general, sin embargo, se vuelven prominentes por primera vez en otoño, cuando las hojas comienzan a perder su clorofila.
Los carotenoides son comunes en muchos seres vivos y dan un color característico a las zanahorias , el maíz , los canarios y los narcisos , así como a las yemas de huevo , los colinabos , los ranúnculos y los plátanos .
Sus brillantes amarillos y naranjas tiñen las hojas de especies de madera dura como nogal , fresno , arce , álamo amarillo , álamo temblón , abedul , cerezo negro , sicómoro , álamo , sasafrás y aliso . Los carotenoides son el pigmento dominante en la coloración de aproximadamente el 15-30% de las especies de árboles. [6]
Antocianinas
Los rojos, los púrpuras y sus combinaciones que decoran el follaje otoñal provienen de otro grupo de pigmentos en las células llamados antocianinas . A diferencia de los carotenoides, estos pigmentos no están presentes en la hoja durante la temporada de crecimiento, pero se producen activamente hacia el final del verano. [6] Se desarrollan a fines del verano en la savia de las células de la hoja, y este desarrollo es el resultado de interacciones complejas de muchas influencias, tanto dentro como fuera de la planta. Su formación depende de la descomposición de los azúcares en presencia de luz brillante a medida que se reduce el nivel de fosfato en la hoja. [10]
Durante la temporada de cultivo de verano, el nivel de fosfato es alto. Tiene un papel vital en la descomposición de los azúcares fabricados por la clorofila, pero en el otoño, el fosfato, junto con otros químicos y nutrientes, se mueve de la hoja al tallo de la planta. Cuando esto sucede, el proceso de descomposición del azúcar cambia, lo que lleva a la producción de pigmentos de antocianina. Cuanto más brillante sea la luz durante este período, mayor será la producción de antocianinas y más brillante será el color resultante. Cuando los días de otoño son brillantes y frescos, y las noches son frías pero no heladas, las coloraciones más brillantes suelen desarrollarse.
Las antocianinas colorean temporalmente los bordes de algunas de las hojas muy jóvenes a medida que se despliegan de los brotes a principios de la primavera. También dan el color familiar a frutas tan comunes como arándanos , manzanas rojas , arándanos , cerezas , fresas y ciruelas .
Las antocianinas están presentes en aproximadamente el 10% de las especies de árboles en las regiones templadas, aunque en ciertas áreas, la más famosa del norte de Nueva Inglaterra, hasta el 70% de las especies de árboles pueden producir el pigmento. [6] En los bosques otoñales, aparecen vívidos en los arces , robles , árboles amargos , liquidámbar , cornejos , tupelos , cerezos y caquis . Estos mismos pigmentos a menudo se combinan con los colores de los carotenoides para crear el naranja más intenso, los rojos ardientes y los bronces típicos de muchas especies de madera dura.
Paredes celulares
El color marrón de las hojas no es el resultado de un pigmento, sino de las paredes celulares, que pueden ser evidentes cuando no se ve ningún pigmento colorante. [6]
Función de los colores otoñales
Tradicionalmente se creía que las plantas de hoja caduca arrojaban sus hojas en otoño principalmente porque los altos costos involucrados en su mantenimiento superarían los beneficios de la fotosíntesis durante el período invernal de poca disponibilidad de luz y temperaturas frías. [11] En muchos casos, esto resultó ser demasiado simplista; otros factores involucrados incluyen la depredación de insectos, [12] la pérdida de agua y los daños causados por fuertes vientos o nevadas.
Las antocianinas, responsables de la coloración rojo-púrpura, se producen activamente en otoño, pero no participan en la caída de las hojas. Se han propuesto varias hipótesis sobre el papel de la producción de pigmentos en la caída de las hojas y, en general, se dividen en dos categorías: interacción con animales y protección frente a factores no biológicos. [6]
Fotoprotección
Según la teoría de la fotoprotección, las antocianinas protegen a la hoja de los efectos nocivos de la luz a bajas temperaturas. [13] [14] Las hojas están a punto de caer, por lo que la protección no es de extrema importancia para el árbol. Sin embargo, la fotooxidación y la fotoinhibición, especialmente a bajas temperaturas, hacen que el proceso de reabsorción de nutrientes sea menos eficiente. Al proteger la hoja con antocianinas, según la teoría de la fotoprotección, el árbol logra reabsorber los nutrientes (especialmente el nitrógeno) de manera más eficiente.
Coevolución
Según la teoría de la coevolución , [15] los colores son señales de advertencia para insectos como los pulgones que utilizan los árboles como hospedadores durante el invierno. Si los colores están relacionados con la cantidad de defensas químicas contra los insectos, los insectos evitarán las hojas rojas y aumentarán su aptitud; al mismo tiempo, los árboles con hojas rojas tienen una ventaja porque reducen su carga parasitaria. Esto se ha demostrado en el caso de los manzanos, donde algunas variedades de manzanas domesticadas, a diferencia de las silvestres , carecen de hojas rojas en otoño. Una mayor proporción de pulgones que evitan los manzanos con hojas rojas logran crecer y desarrollarse en comparación con los que no lo hacen. [dieciséis]Además, existe una compensación entre el tamaño de la fruta, el color de las hojas y la resistencia a los pulgones, ya que las variedades con hojas rojas tienen frutos más pequeños, lo que sugiere un costo para la producción de hojas rojas vinculado a una mayor necesidad de reducir la infestación de pulgones. [dieciséis]
De acuerdo con los árboles de hojas rojas que proporcionan una supervivencia reducida para los pulgones , las especies de árboles con hojas brillantes tienden a seleccionar plagas de áfidos más especializadas que los árboles que carecen de hojas brillantes (los colores otoñales son útiles solo en aquellas especies que coevolucionan con las plagas de insectos en otoño). [17]
La teoría de la coevolución de los colores otoñales fue propuesta por WD Hamilton en 2001 como un ejemplo de la teoría de la señalización evolutiva . [17] Con señales biológicas como las hojas rojas, se argumenta que debido a que son costosas de producir, generalmente son honestas, por lo que señalan la verdadera calidad del comunicador con individuos de baja calidad que no pueden falsificarlas y engañar. Los colores otoñales serían una señal si son costosos de producir o imposibles de falsificar (por ejemplo, si los pigmentos otoñales fueran producidos por la misma vía bioquímica que produce las defensas químicas contra los insectos). [ cita requerida ]
El cambio de color de las hojas antes del otoño también se ha sugerido como adaptaciones que pueden ayudar a socavar el camuflaje de los herbívoros. [18]
Muchas plantas con bayas atraen a las aves con un color de bayas y / o hojas especialmente visible, particularmente rojo brillante. Las aves obtienen una comida, mientras que el arbusto, la vid o, por lo general, un árbol pequeño obtiene las semillas no digeridas que se llevan y se depositan con el estiércol de las aves. La hiedra venenosa es particularmente notable por tener un follaje de color rojo brillante que atrae a las aves hacia sus semillas blanquecinas (que son comestibles para las aves, pero no para la mayoría de los mamíferos).
Alelopatía
El color rojo brillante del otoño de algunas especies de arce se crea mediante procesos distintos de los de la descomposición de la clorofila. Cuando el árbol está luchando por hacer frente a las demandas de energía de una estación cambiante y desafiante, los arces están involucrados en un gasto metabólico adicional para crear antocianinas. Se ha descubierto que estas antocianinas, que crean los tonos rojos visuales, ayudan en la competencia interespecífica al retrasar el crecimiento de los árboles jóvenes cercanos ( alelopatía ). [19]
Turismo
Aunque se produce algo de coloración otoñal dondequiera que se encuentren árboles de hoja caduca, el follaje otoñal de colores más brillantes se encuentra en el hemisferio norte, incluida la mayor parte del sur de Canadá continental , algunas áreas del norte de los Estados Unidos , el norte y oeste de Europa al norte de los Alpes y el Cáucaso. región de Rusia cerca del Mar Negro y Asia Oriental (incluida gran parte del norte y este de China , así como Corea y Japón ). [20] [21]
En el hemisferio sur, se puede observar un colorido follaje otoñal en el sur y centro de Argentina , las regiones sur y sureste de Brasil y el sureste de Australia (incluida Tasmania ). [22]
Influencias climáticas
En comparación con Europa occidental (excluyendo el sur de Europa), América del Norte ofrece muchas más especies de árboles (más de 800 especies y alrededor de 70 robles, en comparación con 51 y tres, respectivamente, en Europa occidental) [23], lo que agrega muchos más colores diferentes al espectáculo. La razón principal es el efecto diferente de las edades de hielo : mientras que en América del Norte, las especies estaban protegidas en las regiones más al sur a lo largo de las montañas que se extienden de norte a sur, este no fue el caso en gran parte de Europa. [24]
El calentamiento global y el aumento de los niveles de dióxido de carbono en la atmósfera pueden retrasar el espectáculo otoñal habitual de colores cambiantes y hojas caídas en los bosques de frondosas del norte en el futuro, y aumentar la productividad forestal. [25] Los experimentos con álamos mostraron que se mantenían más verdes por más tiempo con niveles más altos de CO 2 , independientemente de los cambios de temperatura. [25] Sin embargo, los experimentos durante dos años fueron demasiado breves para indicar cómo los bosques maduros pueden verse afectados con el tiempo. Además, otros factores, como el aumento de los niveles de ozono cerca del suelo ( contaminación por ozono troposférico ), pueden anular los efectos beneficiosos del dióxido de carbono elevado. [26]
Típico follaje de otoño en el país del arce rojo , en las montañas Adirondack del norte del estado de Nueva York
Carril rural inglés en otoño
Diferentes etapas de colores del follaje.
árbol de arce
Finales de otoño con el marrón dominando
Coloración otoñal en Karasawa de las montañas Hodaka en Japón
Coloración otoñal en el parque Katariinapuisto en Lapinjärvi , Finlandia
La ciudad de Stirling, Australia del Sur , ubicada en Adelaide Hills , atrae a muchos turistas durante el otoño.
Una variedad de colores alrededor del templo Tōfuku-ji en Kioto , Japón
Algunos árboles, como este liquidámbar americano en Keokea, Maui , desarrollan colores otoñales llamativos en áreas tropicales o subtropicales.
Referencias
![]() Este artículo incorpora material de dominio público del documento del gobierno de los Estados Unidos : " The USDA Forest Service ".
Este artículo incorpora material de dominio público del documento del gobierno de los Estados Unidos : " The USDA Forest Service ".
- ^ "La ciencia del color en hojas de otoño" . usna.usda.gov . Arboreto Nacional de los Estados Unidos . 6 de octubre de 2011. Archivado desde el original el 11 de enero de 2018 . Consultado el 18 de junio de 2015 .
- ^ Wade, Paul; Arnold, Kathy (16 de septiembre de 2014). "Nueva Inglaterra en el otoño: viaje de su vida - Telegraph" . telegraph.co.uk . The Daily Telegraph . Consultado el 18 de junio de 2015 .
- ^ "BBC - Jardinería - Cómo ser un jardinero - El año de la jardinería - Tema de otoño" . bbc.co.uk . BBC . 17 de septiembre de 2014 . Consultado el 18 de junio de 2015 .
- ^ "Servicio Forestal de Estados Unidos - cuidar la tierra y servir a la gente" . fs.fed.us . Servicio Forestal de los Estados Unidos . 2014 . Consultado el 18 de junio de 2015 .
- ^ "MaineFoliage.com - sitio web oficial de follaje de otoño de Maine" . MaineFoliage.com . MaineFoliage.com. 2013 . Consultado el 18 de junio de 2015 .
- ^ a b c d e f g h Archetti, Marco; Döring, Thomas F .; Hagen, Snorre B .; Hughes, Nicole M .; Cuero, Simon R .; Lee, David W .; Lev-Yadun, Simcha; Manetas, Yiannis; Ougham, Helen J. (2011). "Desentrañar la evolución de los colores del otoño: un enfoque interdisciplinario". Tendencias en Ecología y Evolución . 24 (3): 166–73. doi : 10.1016 / j.tree.2008.10.006 . PMID 19178979 .
- ^ Horie, Y .; Ito, H .; Kusaba, M .; Tanaka, R .; Tanaka, A. (2009). "Participación de la clorofila b reductasa en el paso inicial de la degradación de los complejos de clorofila a / b-proteína de captación de luz en Arabidopsis" . Revista de Química Biológica . 284 (26): 17449–56. doi : 10.1074 / jbc.M109.008912 . PMC 2719385 . PMID 19403948 .
- ^ Zelisko, A .; García-Lorenzo, M .; Jackowski, G .; Jansson, S .; Funk, C. (2005). "AtFtsH6 está involucrado en la degradación del complejo de captación de luz II durante la aclimatación y senescencia a la luz alta" . Actas de la Academia Nacional de Ciencias . 102 (38): 13699–704. Código bibliográfico : 2005PNAS..10213699Z . doi : 10.1073 / pnas.0503472102 . PMC 1224624 . PMID 16157880 .
- ^ Hortensteiner, S. (2006). "Degradación de la clorofila durante la senescencia". Revisión anual de biología vegetal . 57 : 55–77. doi : 10.1146 / annurev.arplant.57.032905.105212 . PMID 16669755 .
- ^ Davies, Kevin M. (2004). Pigmentos vegetales y su manipulación . Wiley-Blackwell . pag. 6. ISBN 978-1-4051-1737-1.
- ^ Thomas, H; Stoddart, JL (1980). "Senescencia de la hoja". Revisión anual de fisiología vegetal . 31 : 83-111. doi : 10.1146 / annurev.pp.31.060180.000503 .
- ^ Labandeira, CC; Dilcher, DL; Davis, DR; Wagner, DL (1994). "Noventa y siete millones de años de asociación angiosperma-insecto: conocimientos paleobiológicos sobre el significado de la coevolución" . Actas de la Academia Nacional de Ciencias . 91 (25): 12278–82. Código bibliográfico : 1994PNAS ... 9112278L . doi : 10.1073 / pnas.91.25.12278 . PMC 45420 . PMID 11607501 .
- ^ Lee, David; Gould, Kevin (2002). "Por qué las hojas se vuelven rojas". Científico estadounidense . 90 (6): 524–531. Código Bibliográfico : 2002AmSci..90..524L . doi : 10.1511 / 2002.6.524 .
- ^ Lee, D; Gould, K (2002). "Antocianinas en hojas y otros órganos vegetativos: una introducción". Avances en la investigación botánica . 37 : 1-16. doi : 10.1016 / S0065-2296 (02) 37040-X . ISBN 978-0-12-005937-9.
- ↑ Archetti, M; Brown, SP (junio de 2004). "La teoría de la coevolución de los colores otoñales" . Proc. Biol. Sci . 271 (1545): 1219–23. doi : 10.1098 / rspb.2004.2728 . PMC 1691721 . PMID 15306345 .
- ↑ a b Archetti, M. (2009). "Evidencia de la domesticación de la manzana para el mantenimiento de los colores otoñales por coevolución" . Actas de la Royal Society B: Ciencias Biológicas . 276 (1667): 2575–80. doi : 10.1098 / rspb.2009.0355 . PMC 2684696 . PMID 19369261 .
- ^ a b Hamilton, WD; Brown, SP (2001). "Los colores de los árboles otoñales como señal de discapacidad" . Actas de la Royal Society B: Ciencias Biológicas . 268 (1475): 1489–93. doi : 10.1098 / rspb.2001.1672 . PMC 1088768 . PMID 11454293 .
- ^ Lev-Yadun, Simcha; Dafni, Amots; Flaishman, Moshe A .; Inbar, Moshe; Izhaki, Ido; Katzir, Gadi; Ne'eman, Gidi (2004). "La coloración de la planta socava el camuflaje de insectos herbívoros". BioEssays . 26 (10): 1126–30. doi : 10.1002 / bies.20112 . PMID 15382135 .
- ^ (Frey & Eldridge, 2005) [ cita requerida ]
- ^ "Alerta de plagas" . Universidad Estatal de Dakota del Sur. 30 de agosto de 1998. Archivado desde el original el 20 de octubre de 2006 . Consultado el 28 de noviembre de 2006 .
- ^ Altman, Daniel (8 de noviembre de 2006). "El follaje de otoño prende fuego a Japón" . Taipei Times . Consultado el 28 de noviembre de 2006 .
- ^ Hutchinson, Carrie (2 de marzo de 2019). "Los 5 mejores lugares de Australia para ver los colores del otoño" . Qantas . Consultado el 22 de julio de 2019 .
- ↑ Heinz Ellenberg , H. Ellenberg: Vegetation Mitteleuropas mit den Alpen: In ökologischer, dynamischer und historischer Sicht, UTB, Stuttgart; Quinta edición, en alemán, ISBN 3-8252-8104-3 , 1996 [ página necesaria ]
- ^ "Botanik online: Pflanzengesellschaften - Sommergrüne Laub- und Mischlaubwälder" (en alemán). Servidor de Biología de la Universidad de Hamburgo. Archivado desde el original el 6 de octubre de 2014 . Consultado el 29 de julio de 2020 .
- ^ a b Taylor, Gail; Tallis, Matthew J .; Giardina, Christian P .; Percy, Kevin E .; Miglietta, Franco; Gupta, Pooja S .; Gioli, Beniamino; Calfapietra, Carlo; Gielen, Birgit (2007). "El futuro CO2 atmosférico conduce a la senescencia otoñal retrasada". Biología del cambio global . 14 (2): 264–75. Código Bibliográfico : 2008GCBio..14..264T . CiteSeerX 10.1.1.384.1142 . doi : 10.1111 / j.1365-2486.2007.01473.x .
- ^ "Los bosques podrían beneficiarse cuando el color del otoño llega tarde" . Newswise . Consultado el 17 de agosto de 2008 .
Lectura adicional
- Guy, Robert D .; Krakowski, Jodie (2003). "Colores de otoño: el lienzo de la naturaleza es una sombrilla de seda" (PDF) . Davidsonia . 14 (4): 111-20. Archivado desde el original (PDF) el 19 de octubre de 2013 . Consultado el 18 de mayo de 2007 .
Enlaces externos
- Tintes otoñales de Henry David Thoreau
- Identificar árboles comunes en otoño por sus colores
- Sanderson, Katharine (2007). "Por qué las hojas de otoño se vuelven rojas". Naturaleza . doi : 10.1038 / news.2007.202 .