
{kind=link}
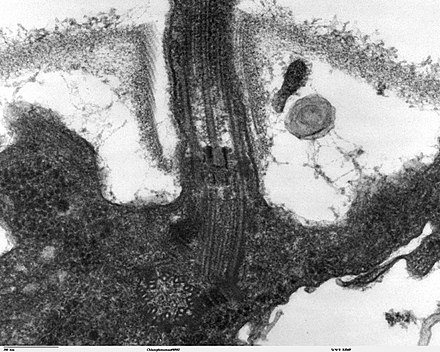
Un cuerpo basal (sinónimo de gránulo basal , cinetosoma y en la literatura citológica más antigua con blefaroplasto ) es una estructura de proteína que se encuentra en la base de un undulipodio eucariota ( cilio o flagelo ). Está formado por un centríolo y varias estructuras proteicas adicionales y es, esencialmente, un centríolo modificado. [1] [2] El cuerpo basal sirve como un sitio de nucleación para el crecimiento de los microtúbulos del axonema . Los centríolos, de los cuales se derivan los cuerpos basales, actúan como sitios de anclaje para las proteínas que a su vez anclan los microtúbulos, y se conocen como el centro organizador de microtúbulos (MTOC). Estos microtúbulos proporcionan estructura y facilitan el movimiento de vesículas y orgánulos dentro de muchas células eucariotas.
Montaje, estructura [ editar ]
Los cilios y los cuerpos basales se forman durante la inactividad o la fase G1 del ciclo celular . Antes de que la célula entre en la fase G1, es decir, antes de la formación del cilio, el centriolo madre actúa como componente del centrosoma .
En las células que están destinadas a tener un solo cilio primario, el centríolo madre se diferencia en el cuerpo basal al entrar en G1 o en reposo. Por tanto, el cuerpo basal de dicha célula se deriva del centríolo. El cuerpo basal se diferencia del centríolo madre en al menos 2 aspectos. Primero, los cuerpos basales tienen pies basales, que están anclados a microtúbulos citoplásmicos y son necesarios para la alineación polarizada del cilio. En segundo lugar, los cuerpos basales tienen fibras de transición en forma de molinete que se originan en los apéndices del centríolo madre. [3]
Sin embargo, en las células multiciliadas, en muchos casos, los cuerpos basales no están hechos de centríolos, sino que se generan de novo a partir de una estructura proteica especial llamada deuterosoma. [4]
Función [ editar ]
Durante la inactividad del ciclo celular, los cuerpos basales organizan los cilios primarios y residen en la corteza celular en las proximidades de la membrana plasmática. Al entrar en el ciclo celular, los cilios se reabsorben y el cuerpo basal migra al núcleo donde funciona para organizar los centrosomas. Los centríolos, los cuerpos basales y los cilios son importantes para la mitosis, la polaridad, la división celular, el tráfico de proteínas, la señalización, la motilidad y la sensación. [5]
Las mutaciones en proteínas que se localizan en los cuerpos basales se asocian con varias enfermedades ciliares humanas, como el síndrome de Bardet-Biedl , [6] el síndrome orofaciodigital , [7] [8] el síndrome de Joubert , [9] la distrofia de conos y bastones , [10] [11 ] Síndrome de Meckel , [12] y nefronoptisis . [13]
La regulación de la producción corporal basal y la orientación espacial es una función del dominio de unión a nucleótidos de la γ-tubulina . [14]
Las plantas carecen de centríolos y solo las plantas inferiores (como musgos y helechos) con espermatozoides móviles tienen flagelos y cuerpos basales. [15]
Referencias [ editar ]
- ^ Jacob M. Schrøder (2011). EB1 y EB3 promueven la biogénesis de los cilios mediante varios mecanismos relacionados con el centrosoma . J Cell Sci 124: 2539-2551; doi: 10.1242 / jcs.085852
- ^ Benjamin Lewin (2007). Celdas . Jones y Bartlett Learning. pag. 359. ISBN 978-0-7637-3905-8. Consultado el 28 de julio de 2019 .
- ^ Sehyun Kim (2013). Montaje de un cilio primario . Opinión actual en biología celular 2013, 25: 506–511
- ^ Deborah A. Klos Dehring (2013). Biogénesis de centriolos mediada por deuterosomas. Celda de desarrollo 27, 103-112
- ^ Chad G. Pearson (2009). Los componentes del cuerpo basal exhiben una dinámica de proteínas diferencial durante el ensamblaje del cuerpo basal naciente . Mol Biol Cell. 1 de febrero; 20 (3): 904–914.
- ^ SJ Ansley, J. Badano, OE Blacque, J. Hill, BE Hoskins, CCLeitch, JC Kim, AJ Ross, ER Eihers, TM Teslovich, et al. La disfunción basal del cuerpo es una causa probable del síndrome de Bardet ‐ Biedl pleiotrópico. Nature, 425 (2003), págs. 628-633.
- ^ MI Ferrante, Z. Zullo, A. Barra, A. Bimonte, N. Messaddeq, M.Studer, P. Dolle, B. Franco. La proteína de tipo I oral-facial-digital es necesaria para la formación de cilios primarios y la especificación del eje izquierdo-derecho. Nat. Genet., 38 (2006), págs. 112-117
- ^ L. Romio, AM Fry, PJ Winyard, S. Malcolm, AS Woolf, SAFeather. OFD1 es una proteína del cuerpo centrosomal / basal expresada durante la transición mesenquimatosa-epitelial en la neurogénesis humana. Mermelada. Soc. Nephrol., 15 (2004), págs. 2566-2568
- ^ HH Arts, D. Doherty, SE van Beersum, MA Parisi, SJ Lettboer, NT Gorden, TA Peters, T. Maerker, K. Voesenek, A. Kartono, H. Ozurek, FMFarin, et al. Las mutaciones en el gen que codifica la proteína corporal basal RPGRIP1L, un interactor nefrocistina-4, causan el síndrome de Joubert. Nat. Genet., 39 (2007), págs. 875-881
- ^ A. Kobayashi, T. Higashide, D. Hamasaki, S. Kubota, H.Sakuma, W. An, T. Tujimaki, MJ McLaren, RG Weleber, G. Inana. La mutación HRG (UNC119) encontrada en la distrofia de conos y bastones causa degeneración de la retina en un modelo transgénico. Invertir. Oftalmol. Vis. Sci., 41 (2000), págs. 3268-3277
- ^ X. Shu, AM Fry, B. Tulloch, JW Manson, JW Crabb, A. Khanna, AJ Faragher, A. Lennon, S. He, P. Trojan. La isoforma RPGR OFR15 co-localiza con RPGRIP1 en centriolos y cuerpos basales e interactúa con nucleofosmina. Tararear. Mol. Genet., 14 (2005), págs. 1193-1197
- ^ M. Kyttala, J. Tallila, R. Salonen, O. Kopra, N. Kohlschmidt, P.Paavola-Sakki, L. Peltonen, M. Kestila. MKS1, que codifica un componente del proteoma del cuerpo basal del aparato flagelar, está mutado en el síndrome de Meckel. Nat. Genet., 38 (2006), págs. 155-157
- ^ YO Winklebauer, JC Schafer, CJ Haycraft, P. Swoboda, BK Yoder. Los homólogos de C. elegans de nefrocistina-1 y nefrocistina-4 son proteínas de la zona de transición de los cilios implicadas en la percepción quimiosensorial. J. Cell Sci., 118 (2005), págs. 5575-5587
- ^ Y. Shang, C.-C. Tsao y MA Gorovsky. 2005. Los análisis mutacionales revelan una función novedosa del dominio de unión a nucleótidos de la gamma-tubulina en la regulación de la biogénesis corporal basal. J. Cell Biol. 171 (6): 1035-44. PMID 16344310
- ^ Philip E. Pack, Ph.D., Notas de Cliff: 4ª edición de Biología AP.
Enlaces externos [ editar ]
- Imagen de histología: 21804loa - Sistema de aprendizaje de histología de la Universidad de Boston - "Ultraestructura de la célula: epitelio ciliado, cilios y cuerpos basales"