
Una escama de pescado es una pequeña placa rígida que surge de la piel de un pez. La piel de la mayoría de los peces está cubierta con estas escamas protectoras , que también pueden proporcionar un camuflaje eficaz mediante el uso de reflejos y coloración , así como posibles ventajas hidrodinámicas. El término escala deriva del francés antiguo "escale", que significa vaina de concha o cáscara. [1]
Las escamas varían enormemente en tamaño, forma, estructura y extensión, desde placas blindadas fuertes y rígidas en peces como camarones y peces cofre , hasta microscópicas o ausentes en peces como anguilas y rapes . La morfología de una escala se puede utilizar para identificar la especie de pez de la que proviene.
La mayoría de los peces óseos están cubiertos con las escamas cicloides del salmón y la carpa , o las escamas ctenoides de la perca , o las escamas ganoides de los esturiones y gars . Los peces cartilaginosos ( tiburones y rayas ) están cubiertos de escamas placoides. En cambio, algunas especies están cubiertas por escudos y otras no tienen una cubierta exterior en parte o toda la piel.
Las escamas de los peces son parte del sistema tegumentario de los peces y se producen a partir de la capa del mesodermo de la dermis , que las distingue de las escamas de los reptiles . [2] [3] Los mismos genes involucrados en el desarrollo de dientes y cabello en mamíferos también están involucrados en el desarrollo de escamas. Las escamas placoides de los peces cartilaginosos también se denominan dentículos dérmicos y son estructuralmente homólogas a los dientes de los vertebrados. La mayoría de los peces también están cubiertos por una capa de moco o limo que puede proteger contra patógenos como bacterias, hongos y virus, y reducir la resistencia de la superficie cuando el pez nada.
Escamas de Thelodont

Se conocen bien las escamas óseas de los thelodontes , la forma más abundante de peces fósiles . Las escamas se formaron y se desprendieron a lo largo de la vida de los organismos y se separaron rápidamente después de su muerte. [4]
El hueso, un tejido que es resistente al daño mecánico y relativamente propenso a la fosilización, a menudo conserva los detalles internos, lo que permite estudiar en detalle la histología y el crecimiento de las escamas. Las escamas comprenden una "corona" que no crece compuesta de dentina , con una superficie superior esmaltada a veces ornamentada y una base de aspidina. [5] Su base de crecimiento está hecha de hueso libre de células, que a veces desarrolló estructuras de anclaje para fijarlo en el costado del pez. [6] Más allá de eso, parece haber cinco tipos de crecimiento óseo, que pueden representar cinco agrupaciones naturales dentro de los thelodonts, o un espectro que varía entre los tejidos meta- (u orto-) dentina y mesodentina de los miembros terminales. [7]Cada uno de los cinco morfos de escala parece parecerse a las escamas de agrupaciones de peces más derivadas, lo que sugiere que los grupos lodontes pueden haber sido grupos de tallos de clados de peces sucesivos. [6]
Sin embargo, usar la morfología de la escala solo para distinguir especies tiene algunos inconvenientes. Dentro de cada organismo, la forma de las escamas varía enormemente según el área del cuerpo, [8] con formas intermedias que aparecen entre diferentes áreas y, para empeorar las cosas, la morfología de las escamas puede que ni siquiera sea constante dentro de un área. Para confundir aún más las cosas, las morfologías de escala no son exclusivas de los taxones y pueden ser indistinguibles en la misma área de dos especies diferentes. [9]
La morfología y la histología de los thelodontes proporciona la herramienta principal para cuantificar su diversidad y distinguir entre especies, aunque, en última instancia, el uso de tales rasgos convergentes es propenso a errores. No obstante, se ha propuesto un marco que comprende tres grupos basándose en la morfología y la histología de la escala. [7] Las comparaciones con las especies modernas de tiburones han demostrado que las escamas de thelodont eran funcionalmente similares a las de los peces cartilaginosos modernos y, de la misma forma, han permitido una amplia comparación entre nichos ecológicos. [10]
Escamas cosmoides
Las escamas cosmoides se encuentran solo en peces antiguos con aletas lobulares , incluidos algunos de los primeros peces pulmonados (subclase Dipnoi ), y en Crossopterygii , incluido el celacanto vivo en una forma modificada (ver escamas elasmoides, más abajo). Probablemente se derivaron de una fusión de escamas placoides. La parte interna de las escamas está hecha de un hueso laminar denso llamado isopedina. Encima de esto se encuentra una capa de hueso esponjoso o vascular provisto de vasos sanguíneos, seguida de una capa compleja similar a la dentina llamada cosmina con una capa externa superficial de vitrodentina . La superficie superior esqueratina . Las escamas cosmoides aumentan de tamaño a través del crecimiento de la capa de hueso laminar. [11]
Escamas elasmoides
Las escamas elasmoides son escamas delgadas e imbricadas compuestas por una capa de hueso de colágeno laminar denso llamado isopedina, por encima de la cual hay una capa de tubérculos generalmente compuestos de hueso, como en Eusthenopteron . La capa de dentina que estaba presente en el primer pez con aletas lobuladas suele estar reducida, como en el celacanto existente , o completamente ausente, como en el pez pulmonado existente y en el Eusthenopteron del Devónico . [12] Las escamas elasmoides han aparecido varias veces a lo largo de la evolución de los peces. Están presentes en algunos peces con aletas lobuladas , como todos los peces pulmonados existentes y algunos extintos , así como los celacantos.que tienen escamas cosmoides modificadas que carecen de cosmina y son más delgadas que las verdaderas escamas cosmoides. También están presentes en algunos tetrapodomorfos como Eusthenopteron , amiidos y teleósteos, cuyas escamas cicloides y ctenoides representan las escamas elasmoides menos mineralizadas.
Las escamas elasmoides de pez cebra se utilizan en el laboratorio para estudiar el proceso de mineralización ósea y se pueden cultivar (mantener) fuera del organismo. [13] [14]
Escamas de Ganoide

Las escamas de ganoides se encuentran en los esturiones , peces espátula , gars , bowfin y bichires . Se derivan de escamas cosmoides y a menudo tienen bordes dentados. Están cubiertos con una capa de dentina dura parecida a un esmalte en lugar de cosmina y una capa de sal ósea inorgánica llamada ganoína en lugar de vitrodentina .
La ganoína es un componente característico de las escamas de ganoides. Es un tejido mineralizado vidrioso, a menudo de varias capas , que cubre las escamas, así como los huesos craneales y los rayos de las aletas en algunos peces con aletas radiadas no teleósteos , [15] como gars , bichires y celacantos . [16] [17] Está compuesto de cristalitos de apatita en forma de varilla . [18] La ganoína es una característica antigua de los peces con aletas radiadas, que se encuentra, por ejemplo, en las escamas del grupo de tallos Cheirolepis actinopteryigian . [17]Aunque a menudo se considera un carácter sinapomórfico de los peces con aletas radiadas, también se encuentran tejidos de ganoína o similares a la ganoína en los acanthodii extintos . [17] Se ha sugerido que la ganoína es homóloga al esmalte dental en los vertebrados [15] o incluso se considera un tipo de esmalte. [18]
Amblypterus striatus | Escamas Ganoides del extinto pez Carbonífero , Amblypterus striatus . (a) muestra la superficie exterior de cuatro de las escamas, y (b) muestra la superficie interior de dos de las escamas. Cada una de las escamas ganoides de forma romboidal de Amblypterus tiene una cresta en la superficie interior que se produce en un extremo en una clavija saliente que encaja en una muesca en la siguiente escala, similar a la forma en que las tejas se pegan juntas en el techo. de una casa. |  |
La mayoría de las escamas ganoides son romboidales (en forma de diamante) y están conectadas por uniones de clavija y encaje. Por lo general, son gruesos y encajan más como una sierra de calar en lugar de superponerse como otras escalas. [19] De esta manera, las escamas de los ganoides son casi impenetrables y son una excelente protección contra la depredación.
El cocodrilo tiene una armadura resistente de escamas de ganoide de forma romboidal . [19]

El esturión tiene filas de escamas ganoides agrandadas en placas de armadura en forma de escudos.
Las escamas ganoides en una aleta de arco son de tamaño reducido y se parecen a las escamas cicloides .

En los esturiones, las escamas se agrandan mucho en placas de armadura a lo largo de los lados y la espalda, mientras que en la aleta de arco las escamas se reducen mucho en grosor para parecerse a las escamas cicloides .
Pendientes hechos con escamas ganoides de un cocodrilo gar
Fósil de un rayfin primitivo con escamas ganoides
Escamas Ganoides en Lepidotes fosilizados , circa. 130 millones de años
Los nativos americanos y la gente del Caribe usaban las duras escamas ganoides del cocodrilo como puntas de flecha, corazas y como escudo para cubrir los arados. En la actualidad la joyería se elabora a partir de estas escalas. [20]
Escamas leptoides
Las escamas leptoides (crestas óseas) se encuentran en peces óseos de orden superior, los teleósteos (el clado más derivado de los peces con aletas radiadas). La parte exterior de estas escamas se abre en abanico con crestas óseas, mientras que la parte interior está cruzada con tejido conectivo fibroso. Las escamas leptoides son más delgadas y más translúcidas que otros tipos de escamas, y carecen de las capas de dentina o esmalte endurecido. A diferencia de las escamas de ganoides, se agregan más escamas en capas concéntricas a medida que el pez crece. [21]
Las escamas leptoides se superponen en una configuración de la cabeza a la cola, como las tejas, lo que las hace más flexibles que las escamas cosmoides y ganoides. Esta disposición permite un flujo de agua más suave sobre el cuerpo y reduce la resistencia . [22] Las escamas de algunas especies exhiben bandas de crecimiento estacional desigual llamadas anillos ( anillo singular ). Estas bandas se pueden utilizar para envejecer los peces .
Las escamas leptoides se presentan en dos formas: cicloide y ctenoide.
Escamas cicloides
Las escamas cicloides (circulares) tienen una textura suave y son uniformes, con un borde o margen exterior liso. Son más comunes en peces con radios de aleta blandas, como el salmón y la carpa .
Los arowana asiáticos tienen grandes escamas cicloides dispuestas sobre el pez en un mosaico de costillas elevadas (izquierda). Las escamas mismas están cubiertas con un delicado patrón de red (derecha). [23] [24] | |
Las escamas cicloides (circulares) se encuentran generalmente en peces parecidos a la carpa o al salmón. |
|

Escamas ctenoideas
Las escamas ctenoideas (dentadas) son como escamas cicloides, excepto que tienen pequeños dientes o espínulas llamadas ctenios a lo largo de sus bordes externos o posteriores. Debido a estos dientes, las escamas tienen una textura rugosa. Por lo general, se encuentran en peces con radios de aleta espinosa, como los peces con forma de perca . Estas escamas casi no contienen hueso y están compuestas por una capa superficial que contiene hidroxiapatita y carbonato de calcio y una capa más profunda compuesta principalmente de colágeno . El esmalte de los otros tipos de escamas se reduce a crestas superficiales y ctenios.
El tamaño de los dientes en las escamas ctenoideas puede variar con la posición, como muestran estas escamas de la cola de rata Cetonurus crassiceps . | |
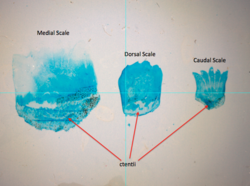

Las escamas ctenoides (dentadas) se encuentran generalmente en peces con forma de perca. |
|




Las escamas ctenoideas, similares a otras estructuras epidérmicas, se originan a partir de placodas y la diferenciación celular distintiva las hace exclusivas de otras estructuras que surgen del tegumento . [26] El desarrollo comienza cerca de la aleta caudal , a lo largo de la línea lateral del pez. [27] El proceso de desarrollo comienza con una acumulación de fibroblastos entre la epidermis y la dermis . [26] Las fibrillas de colágeno comienzan a organizarse en la capa dérmica, lo que conduce al inicio de la mineralización . [26]La circunferencia de las escamas crece primero, seguida del grosor cuando las capas superpuestas se mineralizan juntas. [26]
Las escamas ctenoideas se pueden subdividir en tres tipos:
- Crenate escamas, donde el margen de la escala tiene hendiduras y proyecciones.
- Escamas espinoides , donde la escala tiene espinas que son continuas con la propia escala.
- Verdaderas escamas ctenoides , donde las espinas de la escala son estructuras distintas.
La mayoría de los peces con aletas radiadas tienen escamas ctenoides. Algunas especies de peces planos tienen escamas ctenoides en el lado de los ojos y escamas cicloides en el lado ciego, mientras que otras especies tienen escamas ctenoides en los machos y escamas cicloides en las hembras.
Reflexión

Muchos peces teleósteos están cubiertos con escamas altamente reflectantes que funcionan como pequeños espejos y dan la apariencia de vidrio plateado. La reflexión a través del plateado está muy extendida o es dominante en los peces de mar abierto, especialmente en los que viven en los 100 metros superiores. Se puede lograr un efecto de transparencia plateando para hacer que el cuerpo de un animal sea altamente reflectante. A profundidades medias en el mar, la luz proviene de arriba, por lo que un espejo orientado verticalmente hace que animales como los peces sean invisibles desde el costado. [28]
El pez hacha marino está extremadamente aplanado lateralmente (de lado a lado), dejando el cuerpo de solo milímetros de grosor, y el cuerpo es tan plateado que parece papel de aluminio . Los espejos consisten en estructuras microscópicas similares a las que se utilizan para proporcionar coloración estructural : pilas de entre 5 y 10 cristales de guanina espaciados aproximadamente ¼ de longitud de onda para interferir constructivamente y lograr una reflexión de casi el 100%. En las aguas profundas en las que vive el pez hacha, solo la luz azul con una longitud de onda de 500 nanómetros se filtra hacia abajo y necesita reflejarse, por lo que los espejos separados por 125 nanómetros proporcionan un buen camuflaje. [28]
La mayoría de los peces en la parte superior del océano están camuflados con plateado. En peces como el arenque , que vive en aguas menos profundas, los espejos deben reflejar una mezcla de longitudes de onda y, en consecuencia, el pez tiene pilas de cristales con una variedad de espacios diferentes. Una complicación adicional para los peces con cuerpos redondeados en la sección transversal es que los espejos serían ineficaces si se colocaran planos sobre la piel, ya que no se reflejarían horizontalmente. El efecto de espejo general se logra con muchos reflectores pequeños, todos orientados verticalmente. [28]
Las escamas de pescado con estas propiedades se utilizan en algunos cosméticos, ya que pueden dar un efecto brillante al maquillaje y al lápiz labial. [29]
Escamas placoides
Las escamas placoides (puntiagudas, en forma de diente) se encuentran en los peces cartilaginosos : tiburones , rayas . También se denominan dentículos dérmicos . Las escamas placoides son estructuralmente homólogas a los dientes de los vertebrados ("dentículo" se traduce como "diente pequeño"), y tienen una cavidad pulpar central provista de vasos sanguíneos , rodeada por una capa cónica de dentina , todo lo cual se asienta sobre una placa basal rectangular que descansa sobre la dermis . La capa más externa está compuesta de vitrodentina , un esmalte principalmente inorgánico.-como sustancia. Las escamas placoides no pueden crecer de tamaño, sino que se agregan más escamas a medida que el pez aumenta de tamaño.
También se pueden encontrar escamas similares debajo de la cabeza del arenque dentículo . La cantidad de cobertura de escamas es mucho menor en rayos.
Piel de tiburón
La piel de tiburón está cubierta casi en su totalidad por pequeñas escamas placoides. Las escamas están sostenidas por espinas, que se sienten ásperas cuando se las acaricia hacia atrás, pero cuando se aplanan por el movimiento hacia adelante del agua, crean pequeños vórtices que reducen la resistencia hidrodinámica y reducen la turbulencia , lo que hace que la natación sea más eficiente y silenciosa en comparación con la de los huesos. peces. [30] También desempeña un papel en el antiincrustante al exhibir el efecto de loto . [31]
Todos los dentículos están compuestos por una cavidad pulpar interior con un suministro nervioso y arterial enraizado en la dermis para suministrar moco al dentículo. [32] Los dentículos contienen estructuras de riblet que sobresalen de la superficie de la escala; bajo un microscopio, este riblet puede parecer un gancho o crestas que salen de la escala. La forma general de la protuberancia del dentículo depende del tipo de tiburón y generalmente se puede describir con dos apariencias. [33] La primera es una escala en la que las crestas se colocan lateralmente hacia abajo del tiburón y paralelas al flujo del agua. La segunda forma es una escala lisa con lo que parece un chuletón en forma de gancho que sale de la superficie y apunta hacia el lado posterior del tiburón. [33]Ambas formas de costillas ayudan a crear una capa límite turbulenta que obliga al flujo laminar más lejos de la piel de los tiburones. [34]
A diferencia de los peces óseos, los tiburones tienen un corsé dérmico complicado hecho de fibras colágenas flexibles dispuestas como una red helicoidal que rodea su cuerpo. El corsé funciona como un esqueleto externo, proporcionando sujeción para sus músculos de natación y, por lo tanto, ahorrando energía. [35] Dependiendo de la posición de estas escamas placoides en el cuerpo, pueden ser flexibles y pueden erigirse pasivamente, lo que les permite cambiar su ángulo de ataque. Estas escamas también tienen riblets que están alineados en la dirección del flujo, estos riblets reducen la fuerza de arrastre que actúa sobre la piel del tiburón empujando el vórtice más lejos de la superficie de la piel, inhibiendo cualquier flujo transversal de alta velocidad. [36]
Morfología de la escala
La anatomía general de las escamas varía, pero todas se pueden dividir en tres partes: la corona, el cuello y la base. La flexibilidad de la escala está relacionada con el tamaño de la base de la escala. Las escamas con mayor flexibilidad tienen una base más pequeña y, por lo tanto, están unidas con menos rigidez al estrato laxo. En la corona de los tiburones que nadan rápidamente hay una serie de crestas o crestas paralelas que van desde una dirección anterior a la posterior. [37]
Analizando los tres componentes de la escala se puede concluir que la base del dentículo no entra en contacto con ninguna porción del flujo de fluido. [38] Sin embargo, la corona y el cuello de los dentículos desempeñan un papel clave y son responsables de crear los vórtices turbulentos y los remolinos que se encuentran cerca de la superficie de la piel. [38] Debido al hecho de que los dentículos vienen en tantas formas y tamaños diferentes, se puede esperar que no todas las formas produzcan el mismo tipo de flujo turbulento . Durante un reciente experimento de investigación biomiméticaMuestras de dentículos de tiburón con una microestructura similar a una media luna y probados en un tanque de agua usando una mesa de tracción como un portaobjetos. El experimento mostró que la superficie con dentículos experimentó una reducción de arrastre del 10% en general en comparación con la muestra lisa. La razón de esta reducción de la resistencia se debió al hecho de que los vórtices turbulentos quedaron atrapados entre los dentículos creando una barrera en forma de colchón contra el flujo laminar. [39]Este mismo tipo de experimento fue realizado por otro grupo de investigación que implementó más variación en su muestra biomimética. El segundo grupo llegó a la misma conclusión que el primero, sin embargo, debido a que su experimento contenía más variación dentro de las muestras, pudieron lograr un alto grado de precisión experimental. En conclusión, afirmaron que las formas más prácticas eran más duraderas que las que tenían intrincadas líneas de crestas. Las formas prácticas eran de perfil bajo y contenían una sección transversal en forma de canal trapezoidal o semicircular y eran menos efectivas pero, no obstante, reducían la resistencia en un 6 o 7%. [40]
Reducción de arrastre

Los tiburones reducen la resistencia y el costo total de transporte (COT) a través de múltiples vías diferentes. El arrastre de presión se crea a partir de la diferencia de presión entre el lado anterior y posterior del tiburón debido a la cantidad de volumen que se empuja más allá del tiburón para impulsarse hacia adelante. [41] Este tipo de arrastre también es directamente proporcional al flujo laminar . Cuando el flujo laminar aumenta alrededor del pez, la presión de arrastre también lo hace. [42] El arrastre por fricción es el resultado de la interacción entre el fluido contra la piel del tiburón y puede variar dependiendo de cómo cambia la capa límite contra la superficie del pez. [41]
Los riblets impiden la traslación transversal de los vórtices en sentido de las corrientes en la subcapa viscosa. El mecanismo es complejo y aún no se comprende completamente. Básicamente, los riblets inhiben la formación de vórtices cerca de la superficie porque el vórtice no puede caber en los valles formados por los riblets. Esto empuja el vórtice más hacia arriba desde la superficie, interactuando solo con las puntas de los riblet, sin causar ningún flujo de alta velocidad en los valles. Dado que este flujo de alta velocidad ahora solo interactúa con la punta del riblet, que es un área de superficie muy pequeña, la transferencia de impulso que causa el arrastre ahora es mucho menor que antes, reduciendo así efectivamente el arrastre. Además, esto reduce las fluctuaciones de velocidad transversal, lo que también ayuda en la transferencia de impulso. [37]
Investigaciones recientes han demostrado que existe un régimen previo y posterior a la ruptura en la capa límite cercana a la pared donde la subcapa se espesa a un ritmo decreciente y luego sufre una ruptura abrupta en vórtices turbulentos antes de colapsar finalmente. Este sistema se autorregula por completo y media el ciclo de crecimiento y descomposición; los vórtices se acumulan durante el período de crecimiento y se liquidan abruptamente en formaciones Strouhal de vórtices en forma de horquilla que se levantan de la pared. Los vórtices de elevación son los que empujan la capa límite hacia afuera y lejos de la superficie del tiburón, lo que resulta en una reducción del arrastre general experimentado por los peces. [43]
La textura rugosa, similar al papel de lija , de la piel de tiburón y raya, junto con su dureza, la ha llevado a ser valorada como una fuente de cuero sin curtir , llamado shagreen . Una de las muchas aplicaciones históricas del tiburón tiburón fue la fabricación de empuñaduras para espadas . La textura rugosa de la piel también se usa en la cocina japonesa para hacer ralladores llamados oroshiki , uniendo trozos de piel de tiburón a tablas de madera. El pequeño tamaño de las escamas ralla la comida muy finamente.
Aplicación técnica
En la industria marina, existe un mercado extremadamente grande y la necesidad de superficies antiincrustantes . En términos simples, el ensuciamiento se conoce como el proceso por el cual algo se incrusta con material del entorno circundante, como percebes , algas y lodo verde . Los dentículos dérmicos son un área de investigación extremadamente prometedora para este tipo de aplicación debido al hecho de que los tiburones se encuentran entre los únicos peces sin acumulación o crecimiento en sus escamas. Los estudios de la Marina de los EE. UU. Han demostrado que si se puede diseñar un material biomimético, podría causar un aumento del ahorro de combustible en los buques militares de hasta un 45%. [44]
Hay muchos ejemplos de superficies y materiales biomiméticos basados en la estructura de los organismos acuáticos, incluidos los tiburones. Estas aplicaciones pretenden permitir un movimiento más eficiente a través de medios fluidos como aire, agua y aceite.
También se han utilizado superficies que imitan la piel de los tiburones para evitar que los microorganismos y las algas recubren los cascos de los submarinos y barcos. Una variedad se comercializa como " sharklet ". [45] [46]
Muchos de los nuevos métodos para replicar la piel de tiburón implican el uso de polidimetilsiloxano (PDMS) para crear un molde. Por lo general, el proceso implica tomar un trozo plano de piel de tiburón, cubrirlo con el PDMS para formar un molde y verter PDMS en ese molde nuevamente para obtener una réplica de la piel de tiburón. Este método se ha utilizado para crear una superficie biomimética que tiene propiedades superhidrofóbicas , exhibiendo el efecto de loto . [45] Un estudio encontró que estas superficies biomiméticas redujeron la resistencia hasta en un 9%, [36] mientras que con el movimiento de aleteo la reducción de la resistencia alcanzó el 12,3%. [47]
Junto con las aplicaciones marinas, la industria aeroespacial también puede beneficiarse de estos diseños biomiméticos. El modelado paramétrico se ha realizado en dentículos de tiburón con una amplia gama de variaciones de diseño, como generadores de vórtices de perfil alto y bajo. [48] Estos modelos biomiméticos fueron diseñados y analizados para ver los efectos de la aplicación de estructuras similares a dentículos a las alas de varios aviones. Durante la simulación, se observó que la muestra alteraba la forma en que los ángulos de ataque bajo y altoreaccionó. De las muestras de perfil bajo y alto probadas, los generadores de vórtice de perfil bajo superaron las actuales estructuras de ala suave en un 323%. Este aumento en el rendimiento se debe a una burbuja de separación en la estela de los dentículos y a los vórtices que reponen el impulso perdido en la capa límite debido a la fricción de la piel. [48]
Escudos

Los escudos son similares a las escalas y tienen la misma función. A diferencia de las escamas de los peces, que se forman a partir de la epidermis, los escudos se forman en la capa vascular inferior de la piel y el elemento epidérmico es solo la superficie superior. Al formarse en la dermis viva, los escudos producen una capa externa córnea, que es superficialmente similar a la de las escamas.
Scute viene del latín para escudo y puede tomar la forma de:
- una placa ósea externa en forma de escudo, o
- una escama modificada y engrosada que a menudo tiene quillas o espinas, o
- una escama proyectada, modificada (rugosa y fuertemente estriada), generalmente asociada con la línea lateral, o en el pedúnculo caudal formando quillas caudales, oa lo largo del perfil ventral.
Algunos peces, como el pez piña , están cubiertos total o parcialmente de escudos. Los arenques de río y los aletas roscadas tienen una fila abdominal de escudos, que son escamas con puntas elevadas y afiladas que se utilizan para protección. Algunos gatos tienen una fila de escudos siguiendo la línea lateral a cada lado.
Desarrollo de escala
Las escamas suelen aparecer tarde en el desarrollo de los peces. En el caso del pez cebra , pasan 30 días después de la fertilización antes de que las diferentes capas necesarias para comenzar a formar las escamas se hayan diferenciado y organizado. Para ello es necesario que se produzca la consolidación del mesénquima , luego se induzca la morfogénesis y finalmente se produzca el proceso de diferenciación o metamorfosis tardía . [49] [50]
- Consolidación del mesénquima: La consolidación o estructuración del mesénquima se origina durante el desarrollo de la dermis . Este proceso depende de si el pez es cartilaginoso u óseo. Para los peces cartilaginosos, la estructuración se origina a través de la formación de dos capas. El primero es superficial y ancho y el segundo es delgado y compacto. Estas dos capas están separadas por células mesenquimales . Los peces óseos generan un sustrato acelular organizado perpendicularmente por fibras de colágeno . Posteriormente, para ambos peces los fibroblastos se alargan. Estos penetran en la capa compacta del mesénquima, que se consolida antes de la formación de la escala, para iniciar la placa dérmica. [49][50] [51]
- Inducción de la morfogénesis: La morfogénesis se debe a la formación de la papila epidérmica , que se genera al unir la epidermis y la dermis mediante un proceso de invaginación . La morfogénesis comienza en el momento en que los fibroblastos se trasladan a la parte superior del mesénquima compacto. A lo largo de este proceso, las células basales del epitelio forman una capa delimitante, que se ubica en la parte superior del mesénquima. Posteriormente, estas células se diferenciarán en la zona donde surgirá el primordio de escamas . [49] [50] [51]
- Diferenciación o metamorfosis tardía: Esta diferenciación se genera mediante dos formas diferentes según el tipo de escala que se esté formando. La formación de escamas elasmoides (cicloides y ctenoides) se produce mediante la formación de un espacio entre la matriz de la papila epidérmica. Este espacio contiene fibras de colágeno. Alrededor de este espacio se diferencian los elasmoblastos y son los encargados de generar el material necesario para la formación de la escala. Posteriormente, se produce la mineralización de la matriz , lo que permite que las escamas adquieran la característica rígida que las identifica. [49] [50] [51]
A diferencia de las escamas elasmoides, las escamas ganoides están compuestas de colágeno mineralizado y no mineralizado en diferentes regiones. La formación de estos se produce por la entrada de las células superficiales del mesénquima en la matriz, esta última está compuesta por fibras de colágeno y se localiza alrededor de los capilares vasculares, dando lugar así a cavidades vasculares. En este punto, los elasmoblastos son reemplazados por osteoblastos , formando así hueso. Los parches de la matriz de la escama que no están osificados están compuestos por colágeno compactado que le permiten mantener la unión con el mesénquima. Esto se conoce como fibras de Sharpey . [49] [50] [51]
Uno de los genes que regula el desarrollo de la formación de escamas en los peces es el gen del erizo sónico (shh), que mediante la proteína (shh), involucrada en la organogénesis y en el proceso de comunicación celular , posibilita la formación de las escamas. [52] [53] La apolipoproteína E (ApoE), que permite el transporte y metabolismo de los triglicéridos y el colesterol , tiene una interacción con shh, porque ApoE proporciona colesterol a la vía de señalización shh . Se ha demostrado que durante el proceso de diferenciación e interacción celular, el nivel de transcripción de ApoE es alto, lo que ha llevado a la conclusión de que esta proteína es importante para el desarrollo tardío de escamas. [52] [53]
Escalas modificadas

Los diferentes grupos de peces han desarrollado una serie de escamas modificadas para cumplir diversas funciones.
- Casi todos los peces tienen una línea lateral , un sistema de mecanorreceptores que detectan los movimientos del agua. En los peces óseos, las escamas a lo largo de la línea lateral tienen poros centrales que permiten que el agua entre en contacto con las células sensoriales.
- Las espinas de la aleta dorsal de los tiburones cazón y las quimeras, las espinas de la cola de las mantarrayas y los dientes de "sierra" de los peces sierra y los tiburones sierra son escamas placoides fusionadas y modificadas.
- El pez cirujano tiene una hoja en forma de bisturí, que es una escala modificada, a cada lado del pedúnculo caudal . [54]
- Algunos arenques , anchoas y semipicos tienen escamas caducas , que se desprenden fácilmente y ayudan a escapar de los depredadores.
- Los dardos machos de Percina tienen una fila de escamas caducas agrandadas entre las aletas pélvicas y el ano .
- Los peces puercoespín tienen escamas modificadas en grandes espinas externas .
- Por el contrario, el pez globo tiene espinas más delgadas y ocultas que los peces puercoespín, que se vuelven visibles solo cuando el pez se hincha. A diferencia del pez puercoespín, estas espinas no son escamas modificadas, sino que se desarrollan bajo el control de la misma red de genes que producen plumas y pelos en otros vertebrados. [55] [56]
Los peces puercoespín tienen escamas modificadas en espinas .
Las espinas del pez globo no son escamas modificadas, sino que están desarrolladas por una red genética independiente.

Peces sin escamas
El pez mandarín carece de escamas y se protege con una capa de limo amargo y maloliente.

Los peces sin escamas suelen desarrollar alternativas a la protección que las escamas pueden proporcionar, como la piel dura y curtida o las placas óseas.
- Los peces sin mandíbulas ( lampreas y brujas ) tienen la piel lisa, sin escamas y sin hueso dérmico. [57] Las lampreas obtienen cierta protección de una piel dura y curtida. Los peces bruja exudan grandes cantidades de limo o mucosidad si están amenazados. [58] Pueden atarse con un nudo simple , raspando la baba a medida que avanzan y liberándose de un depredador. [59]
- La mayoría de las anguilas no tienen escamas, aunque algunas especies están cubiertas de pequeñas escamas cicloides lisas.
- La mayoría de los bagres carecen de escamas, aunque varias familias tienen chalecos antibalas en forma de placas dérmicas o algún tipo de escudo. [60]
- El pez mandarín carece de escamas y tiene una capa de limo amargo y maloliente que bloquea las enfermedades y probablemente desalienta a los depredadores, lo que implica que su coloración brillante es aposemática . [61]
- El rape tiene la piel flácida y delgada, a menudo cubierta con espinas o tubérculos dérmicos bifurcados , pero no tienen escamas regulares. Dependen del camuflaje para evitar la atención de los depredadores, mientras que su piel suelta dificulta que los depredadores los agarren.
Muchos grupos de peces óseos, incluidos peces pipa , caballitos de mar , peces cofre , cazadores furtivos y varias familias de espinosos , han desarrollado placas óseas externas, estructuralmente parecidas a escamas placoides, como armadura protectora contra los depredadores.
- Los caballitos de mar carecen de escamas pero tienen la piel fina estirada sobre una armadura de placas óseas dispuestas en anillos a lo largo de sus cuerpos.
- En el pez cofre, las placas se fusionan para formar una concha rígida o exoesqueleto que encierra todo el cuerpo. Estas placas óseas no son escamas modificadas, sino piel osificada. Debido a esta armadura pesada, el pez cofre se limita a movimientos lentos, pero pocos otros peces pueden comerse a los adultos.
Las anguilas parecen sin escamas, pero algunas especies están cubiertas de pequeñas escamas cicloides lisas. | |
Los peces cofre tienen placas de piel osificada fusionadas para formar una concha rígida.
Los caballitos de mar tienen la piel fina estirada sobre placas óseas dispuestas en anillos.
Algunos peces, como el hoki y el pez espada, nacen con escamas, pero las arrojan a medida que crecen.
Los filetes tienen escamas rugosas que no se superponen con pequeños picos, por lo que se les llama filefish. Algunos peces lima parecen sin escamas porque sus escamas son muy pequeñas.
Aparecen escamas prominentes en el atún solo a lo largo de la línea lateral y en el corsé , una banda protectora de escamas engrosadas y agrandadas en la región del hombro. En la mayor parte de su cuerpo, los atunes tienen escamas tan pequeñas que, a una inspección casual, parecen sin escamas. [62]
Algunos peces lima parecen sin escamas porque sus escamas son muy pequeñas.
Para un examen casual, el atún parece en gran parte libre de escamas, pero no lo es.
Levítico
Un pasaje célebre en Levítico declara que, "de todo lo que hay en las aguas ... en los mares y en los ríos" los que no tienen aletas ni escamas "os serán abominación" y no se podrán comer. . [63] Esto elimina todos los invertebrados acuáticos como abominaciones e inmundos , así como cualquier pez que carezca de escamas (no parece haber peces que carezcan de aletas).
Según el chok o decretos divinos de la Torá y el Talmud , para que un pez sea declarado kosher , debe tener escamas y aletas. [64] La definición de "escama" difiere de las definiciones presentadas en biología en que las escamas de un pez kosher deben ser visibles a simple vista y pueden quitarse fácilmente de la piel con la mano o con un cuchillo. [64] Según la agencia de certificación kosher de la Unión Ortodoxa , un pez es kosher si las escamas se pueden quitar sin rasgar su piel. [sesenta y cinco]Por lo tanto, la carpa y el salmón son kosher, mientras que un tiburón, cuyas escamas son microscópicas, un esturión, cuyos escudos no se pueden quitar fácilmente sin cortarlos del cuerpo, no son todos kosher. Otros peces no kosher incluyen el bagre, las anguilas, el bacalao de agua dulce , la caballa serpiente y el pez globo. [64]
Lepidofagia

La lepidofagia (griego antiguo para comer escamas ) es un comportamiento de alimentación especializado en peces que implica comer escamas de otros peces. [67] La lepidofagia ha evolucionado de forma independiente en al menos cinco familias de agua dulce y siete familias marinas. [68]
Las escamas de pescado pueden ser nutritivas y contienen una porción dérmica y una capa de moco rico en proteínas además de las capas de queratina y esmalte . Son una rica fuente de fosfato de calcio . [68] Sin embargo, la energía gastada para hacer un ataque frente a la cantidad de escamas consumidas por ataque pone un límite al tamaño de los peces lepidófagos, y por lo general son mucho más pequeños que sus presas . [68] El comportamiento alimentario de las escamas generalmente evoluciona debido a la falta de alimentos y las condiciones ambientales extremas. Comer escamas y la piel que las rodea proporciona nutrientes ricos en proteínas que pueden no estar disponibles en otros lugares del nicho. [69]
Las mandíbulas de los peces normalmente muestran simetría bilateral . Se produce una excepción con el cíclido que se alimenta de escamas Perissodus microlepis . Las mandíbulas de este pez se presentan en dos formas morfológicas distintas . Un morph tiene su mandíbula torcida hacia la izquierda, lo que le permite comer escamas más fácilmente en el flanco derecho de su víctima. El otro morfo tiene su mandíbula torcida hacia la derecha, lo que facilita comer escamas en el flanco izquierdo de su víctima. La abundancia relativa de los dos morfos en las poblaciones está regulada por la selección dependiente de la frecuencia . [66] [70] [71]
Ver también
- Determinación de la edad en peces
- Coloración animal
- Reflectores de animales
- Cristales fotónicos
- Escama de reptil
- Escala (zoología)
- Armadura de escamas
- Escamas de serpiente
- Urokotori - escalador de pescado japonés
Referencias
- ^ Escala Etymonline . Consultado el 28 de abril de 2019.
- ^ Mongera, A .; Nüsslein-Volhard, C. (2013). "Las escamas de pescado surgen del mesodermo" . Biología actual . 23 (9): R338 – R339. doi : 10.1016 / j.cub.2013.02.056 . PMID 23660349 .
- ^ Sharpe, PT (2001). "Desarrollo de escamas de pescado: ¿Cabello hoy, dientes y escamas ayer?" . Biología actual . 11 (18): R751 – R752. doi : 10.1016 / S0960-9822 (01) 00438-9 . PMID 11566120 . S2CID 18868124 .
- ^ Turner, S .; Tarling, DH (1982). "Thelodont y otras distribuciones agnathan como pruebas de reconstrucciones continentales del Paleozoico Inferior". Paleogeografía, Paleoclimatología, Paleoecología . 39 (3–4): 295–311. Código bibliográfico : 1982PPP .... 39..295T . doi : 10.1016 / 0031-0182 (82) 90027-X .
- ^ Märss, T. (2006). "Ultraescultura exoesquelética de vertebrados tempranos". Revista de Paleontología de Vertebrados . 26 (2): 235–252. doi : 10.1671 / 0272-4634 (2006) 26 [235: EUOEV] 2.0.CO; 2 .
- ↑ a b Janvier, Philippe (1998). "Los primeros vertebrados y sus parientes existentes". Vertebrados tempranos . Prensa de la Universidad de Oxford . págs. 123-127. ISBN 978-0-19-854047-2.
- ↑ a b Turner, S. (1991). "Monofilia e interrelaciones de los Thelodonti". En MM Chang; YH Liu; GR Zhang (eds.). Vertebrados tempranos y problemas relacionados de la biología evolutiva . Science Press, Beijing. págs. 87-119.
- ^ Märss, T. (1986). "Escuamación del thelodont agnathan Phlebolepis ". Revista de Paleontología de Vertebrados . 6 (1): 1–11. doi : 10.1080 / 02724634.1986.10011593 .
- ^ Botella, H .; JI Valenzuela-Ríos; P. Carls (2006). "Un nuevo telodonto del Devónico temprano de Celtiberia (España), con una revisión de los telodontos españoles". Paleontología . 49 (1): 141-154. doi : 10.1111 / j.1475-4983.2005.00534.x .
- ↑ Ferrón, Humberto G .; Botella, Héctor (2017). "Escuamación y ecología de thelodonts" . PLOS ONE . 12 (2): e0172781. Código Bibliográfico : 2017PLoSO..1272781F . doi : 10.1371 / journal.pone.0172781 . PMC 5328365 . PMID 28241029 .
- ^ MICHAEL ALLABY "escala cosmoide". Diccionario de zoología. . Encyclopedia.com. 29 de octubre de 2019 < https://www.encyclopedia.com >
- ^ Zylberberg, L., Meunier, FJ, Laurin, M. (2010). Un estudio microanatómico e histológico del esqueleto dérmico poscraneal en el sarcopterigio del Devónico Eusthenopteron foordi , Acta Palaeontologica Polonica 55: 459–470.
- ^ Bergen, Dylan JM; Kague, Erika; Hammond, Chrissy L. (2019). "El pez cebra como un modelo emergente para la osteoporosis: una plataforma de prueba primaria para la detección de nuevos compuestos osteoactivos" . Fronteras en endocrinología . 10 : 6. doi : 10.3389 / fendo.2019.00006 . ISSN 1664-2392 . PMC 6361756 . PMID 30761080 .
- ↑ de Vrieze, E .; van Kessel, MAHJ; Peters, HM; Spanings, FAT; Flik, G .; Metz, JR (1 de febrero de 2014). "La prednisolona induce un fenotipo similar a la osteoporosis en la regeneración de escamas de pez cebra". Osteoporosis International . 25 (2): 567–578. doi : 10.1007 / s00198-013-2441-3 . ISSN 1433-2965 . PMID 23903952 . S2CID 21829206 .
- ↑ a b Zylberberg, L .; Sire, J. -Y .; Nanci, A. (1997). "Inmunodetección de proteínas similares a la amelogenina en la ganoína de escamas de regeneración experimental de Calamoichthys calabaricus, un pez actinopterigio primitivo" . El registro anatómico . 249 (1): 86–95. doi : 10.1002 / (SICI) 1097-0185 (199709) 249: 1 <86 :: AID-AR11> 3.0.CO; 2-X . PMID 9294653 .
- ↑ Sire, Jean-Yves; Donoghue, Philip CJ; Vickaryous, Matthews K. (2009). "Origen y evolución del esqueleto tegumentario en vertebrados no tetrápodos" . Revista de anatomía . 214 (4): 409–440. doi : 10.1111 / j.1469-7580.2009.01046.x . ISSN 0021-8782 . PMC 2736117 . PMID 19422423 .
- ↑ a b c Richter, M. (1995). "Un estudio microestructural del tejido ganoine de vertebrados inferiores seleccionados". Revista Zoológica de la Sociedad Linneana . 114 (2): 173–212. doi : 10.1006 / zjls.1995.0023 .
- ^ a b Bruet, BJF; Song, J .; Boyce, MC; Ortiz, C. (2008). "Principios de diseño de materiales de armaduras de peces antiguos". Materiales de la naturaleza . 7 (9): 748–756. Código Bibliográfico : 2008NatMa ... 7..748B . doi : 10.1038 / nmat2231 . PMID 18660814 .
- ↑ a b Sherman, Vincent R .; Yaraghi, Nicholas A .; Kisailus, David; Meyers, Marc A. (1 de diciembre de 2016). "Influencias microestructurales y geométricas en las escamas protectoras de la espátula Atractosteus" . Revista de la interfaz de la Royal Society . 13 (125): 20160595. doi : 10.1098 / rsif.2016.0595 . ISSN 1742-5689 . PMC 5221522 . PMID 27974575 .
- ^ "Plan de restauración y gestión de Missouri Alligator Gar" (PDF) . División de Pesca del Departamento de Conservación de Missouri. 22 de enero de 2013. Archivado desde el original (PDF) el 6 de mayo de 2016 . Consultado el 12 de abril de 2019 .
- ^ Lagler, KF, JE Bardach y RR Miller (1962) Ictiología . Nueva York: John Wiley & Sons.
- ^ Ballard, Bonnie; Cheek, Ryan (2 de julio de 2016). Medicina de animales exóticos para el técnico veterinario . John Wiley e hijos. ISBN 978-1-118-92421-1.
- ^ Pouyaud, L .; Sudarto, Guy G. Teugels (2003). "Las diferentes variedades de color de la arowana asiática Scleropages formosus (Osteoglossidae) son especies distintas: evidencias morfológicas y genéticas". Cybium . 27 (4): 287-305.
- ^ Ismail, M. (1989). Sistemática, Zoogeografía y Conservación de los Peces de Agua Dulce de Malasia Peninsular (Ed. Tesis Doctoral). Universidad Estatal de Colorado.
- ^ EJ Brill (1953). Los peces del archipiélago indoaustraliano . EJ Brill. págs. 306-307.
- ^ a b c d Kawasaki, Kenta C., "Un análisis genético de la morfología de la escala de cíclidos" (2016). Tesis de Maestría Mayo de 2014 - actualidad. 425. http://scholarworks.umass.edu/masters_theses_2/425
- ^ Helfman, Gene (2009). La diversidad de la biología, la evolución y la ecología de los peces . Wiley-Blackwell.
- ↑ a b c Herring, Peter (2002). La biología del océano profundo . Oxford: Prensa de la Universidad de Oxford. págs. 193-195. ISBN 9780198549567.
- ^ "Probablemente hay escamas de pescado en tu lápiz labial" . HuffPost India . 23 de abril de 2015 . Consultado el 6 de mayo de 2019 .
- ^ Martín, R. Aidan. "Piel de los dientes" . Consultado el 28 de agosto de 2007 .
- ^ Fürstner, Reiner; Barthlott, Wilhelm; Neinhuis, Christoph; Walzel, Peter (1 de febrero de 2005). "Propiedades humectantes y autolimpiantes de superficies artificiales superhidrofóbicas". Langmuir . 21 (3): 956–961. doi : 10.1021 / la0401011 . ISSN 0743-7463 . PMID 15667174 .
- ↑ Lauder, George V .; Wainwright, Dylan K .; Domel, August G .; Weaver, James C .; Wen, Li; Bertoldi, Katia (2016). "Estructura, biomimética y dinámica de fluidos de las superficies de piel de pescado" . Fluidos de revisión física . 1 (6): 060502. Código bibliográfico : 2016PhRvF ... 1f0502L . doi : 10.1103 / PhysRevFluids.1.060502 . S2CID 18118663 .
- ^ a b Feld, Katrine; Kolborg, Anne Noer; Nyborg, Camilla Marie; Salewski, Mirko; Steffensen, John Fleng; Berg-Sørensen, Kirstine (24 de mayo de 2019). "Dentículos dérmicos de tres especies de tiburones que nadan lentamente: microscopía y visualización de flujo" . Biomiméticos . 4 (2): 38. doi : 10.3390 / biomimetics4020038 . ISSN 2313-7673 . PMC 6631580 . PMID 31137624 .
- ^ Fletcher, Thomas; Altringham, John; Peakall, Jeffrey; Wignall, Paul; Dorrell, Robert (7 de agosto de 2014). "Hidrodinámica de peces fósiles" . Actas de la Royal Society B: Ciencias Biológicas . 281 (1788): 20140703. doi : 10.1098 / rspb.2014.0703 . ISSN 0962-8452 . PMC 4083790 . PMID 24943377 .
- ^ Martín, R. Aidan. "La importancia de ser cartilaginoso" . Centro ReefQuest para la Investigación de Tiburones . Consultado el 29 de agosto de 2009 .
- ^ a b Hage, W .; Bruse, M .; Bechert, DW (1 de mayo de 2000). "Experimentos con riblets tridimensionales como modelo idealizado de piel de tiburón". Experimentos en fluidos . 28 (5): 403–412. Código Bibliográfico : 2000ExFl ... 28..403B . doi : 10.1007 / s003480050400 . ISSN 1432-1114 . S2CID 122574419 .
- ^ a b Motta, Philip; Habegger, Maria Laura; Lang, Amy; Hueter, Robert; Davis, Jessica (1 de octubre de 2012). "Escala de morfología y flexibilidad en el marrajo dientuso Isurus oxyrinchus y el tiburón puntas negras Carcharhinus limbatus". Revista de morfología . 273 (10): 1096-1110. doi : 10.1002 / jmor.20047 . ISSN 1097-4687 . PMID 22730019 . S2CID 23881820 .
- ^ a b Dou, Zhaoliang; Wang, Jiadao; Chen, Darong (1 de diciembre de 2012). "Investigación biónica sobre escamas de pescado para la reducción de la resistencia" . Revista de Ingeniería Biónica . 9 (4): 457–464. doi : 10.1016 / S1672-6529 (11) 60140-6 . ISSN 1672-6529 . S2CID 137143652 .
- ^ "Investigaciones experimentales sobre las características de reducción de arrastre de la superficie biónica con microestructuras de escamas de peces que atrapan el agua" (PDF) .
- ^ Palmer, Colin; Young, Mark T. (14 de enero de 2015). "Reducción de arrastre superficial y control de separación de flujo en vertebrados pelágicos, con implicaciones para la interpretación de morfologías de escala en taxones fósiles" . Ciencia Abierta de la Royal Society . 2 (1): 140163. Bibcode : 2015RSOS .... 240163P . doi : 10.1098 / rsos.140163 . ISSN 2054-5703 . PMC 4448786 . PMID 26064576 .
- ↑ a b Lauder, George V .; Wainwright, Dylan K .; Domel, August G .; Weaver, James C .; Wen, Li; Bertoldi, Katia (18 de octubre de 2016). "Estructura, biomimética y dinámica de fluidos de las superficies de piel de pescado" . Fluidos de revisión física . 1 (6): 060502. Código bibliográfico : 2016PhRvF ... 1f0502L . doi : 10.1103 / PhysRevFluids.1.060502 .
- ^ Muthuramalingam, Muthukumar; Villemin, Leo S .; Bruecker, Christoph (29 de abril de 2019). "Formación de rayas en el flujo sobre matrices de escala de peces biomiméticos". La Revista de Biología Experimental . 222 (Pt 16): jeb205963. arXiv : 1904.12752 . Código bibliográfico : 2019arXiv190412752M . doi : 10.1242 / jeb.205963 . PMID 31375542 . S2CID 139103148 .
- ^ Bandyopadhyay, Promode R .; Hellum, Aren M. (23 de octubre de 2014). "Modelado de cómo los patrones de piel de tiburón y delfín controlan los patrones de vorticidad de turbulencia de pared de transición utilizando mecanismos de restablecimiento de fase espacio-temporal" . Informes científicos . 4 : 6650. Código Bibliográfico : 2014NatSR ... 4E6650B . doi : 10.1038 / srep06650 . ISSN 2045-2322 . PMC 4206846 . PMID 25338940 .
- ^ Magin, Chelsea M .; Cooper, Scott P .; Brennan, Anthony B. (1 de abril de 2010). "Estrategias antiincrustantes no tóxicas" . Materiales hoy . 13 (4): 36–44. doi : 10.1016 / S1369-7021 (10) 70058-4 . ISSN 1369-7021 .
- ^ a b Liu, Yunhong; Li, Guangji (15 de diciembre de 2012). "Un nuevo método para producir el" Efecto Lotus "en la piel de un tiburón biomimético". Revista de ciencia coloide y de interfaz . 388 (1): 235–242. Código Bib : 2012JCIS..388..235L . doi : 10.1016 / j.jcis.2012.08.033 . ISSN 0021-9797 . PMID 22995249 .
- ^ "Descubrimiento de Sharklet | Sharklet Technologies, Inc" . www.sharklet.com . Consultado el 26 de septiembre de 2018 .
- ↑ Lauder, George V .; Oeffner, Johannes (1 de marzo de 2012). "La función hidrodinámica de la piel de tiburón y dos aplicaciones biomiméticas" . Revista de Biología Experimental . 215 (5): 785–795. doi : 10.1242 / jeb.063040 . ISSN 1477-9145 . PMID 22323201 .
- ↑ a b Domel, August G .; Saadat, Mehdi; Weaver, James C .; Haj-Hariri, Hossein; Bertoldi, Katia; Lauder, George V. (28 de febrero de 2018). "Diseños inspirados en la piel de tiburón que mejoran el rendimiento aerodinámico" . Revista de la interfaz de la Royal Society . 15 (139): 20170828. doi : 10.1098 / rsif.2017.0828 . PMC 5832729 . PMID 29436512 .
- ^ a b c d e Sire, JY; Huysseune, ANN (2003). "Formación de tejidos dérmicos esqueléticos y dentales en peces: un enfoque comparativo y evolutivo". Revisiones biológicas . 78 (2): 219–249. doi : 10.1017 / S1464793102006073 . PMID 12803422 . S2CID 19556201 .
- ↑ a b c d e Le Guellec, D .; Morvan-Dubois, G .; Padre, JY (2004). "Desarrollo de la piel en peces óseos con especial énfasis en la deposición de colágeno en la dermis del pez cebra ( Danio rerio )" . Revista Internacional de Biología del Desarrollo . 48 (2-3): 217-231. doi : 10.1387 / ijdb.15272388 . PMID 15272388 .
- ↑ a b c d Sire, JY (2001). "Dientes fuera de la boca en peces teleósteos: cómo beneficiarse de un accidente de desarrollo". Evolución y desarrollo . 3 (2): 104–108. doi : 10.1046 / j.1525-142x.2001.003002104.x . PMID 11341672 . S2CID 13353402 .
- ^ a b Sire, JY; Akimenko, MA (2003). "Desarrollo de escala en peces: una revisión, con descripción de la expresión de erizo sónico (shh) en el pez cebra ( Danio rerio )" . Revista Internacional de Biología del Desarrollo . 48 (2–3): 233–247. doi : 10.1387 / ijdb.15272389 . PMID 15272389 .
- ^ a b Monnot, MJ; Babin, PJ; Poleo, G .; Andre, M .; Laforest, L .; Ballagny, C .; Akimenko, MA (1999). "Expresión epidérmica del gen de la apolipoproteína E durante el desarrollo de aletas y escamas y la regeneración de aletas en pez cebra" . Dinámica del desarrollo . 214 (3): 207–215. doi : 10.1002 / (SICI) 1097-0177 (199903) 214: 3 <207 :: AID-AJA4> 3.0.CO; 2-5 . PMID 10090147 .
- ^ Sorenson, L .; Santini, F .; Carnevale, G .; Alfaro, ME (2013). "Un árbol de tiempo de múltiples locus de peces cirujano (Acanthuridae, Percomorpha), con taxonomía familiar revisada". Filogenética molecular y evolución . 68 (1): 150-160. doi : 10.1016 / j.ympev.2013.03.014 . PMID 23542000 .
- ^ Cómo el pez globo obtuvo sus espinas locas Phys.org , 25 de julio de 2019.
- ^ Shono, T .; Thiery, AP; Cooper, RL; Kurokawa, D .; Britz, R .; Okabe, M .; Fraser, GJ (2019). "Evolución y diversidad de desarrollo de las espinas de la piel en peces globo" . iScience . 19 : 1248-1259. doi : 10.1016 / j.isci.2019.06.003 . PMC 6831732 . PMID 31353167 .
- ^ Coolidge E, Hedrick MS y Milsom WK (2011) "Sistemas de ventilación" . En: McKenzie DJ, Farrell AP y Brauner CJ (Eds) Fisiología de peces: peces primitivos , Elsevier, páginas 182-213. ISBN 9780080549521
- ^ Rothschild, Anna (1 de abril de 2013). "Baba de pez bruja: ¿La ropa del futuro?" . BBC News . Consultado el 2 de abril de 2013 .
- ^ Yong, Ed (23 de enero de 2019). "Nadie está preparado para baba de pez bruja" . El Atlántico . Consultado el 26 de enero de 2019 .
- ^ Friel, JP; Lundberg, JG (1996). " Micromyzon akamai , gen. Et sp. Nov., Un bagre banjo pequeño y sin ojos (Siluriformes: Aspredinidae) de los canales de los ríos de la cuenca baja del Amazonas". Copeia . 1996 (3): 641–648. doi : 10.2307 / 1447528 . JSTOR 1447528 .
- ^ Sadovy, Y .; Randall, JE; Rasotto, Maria B. (mayo de 2005). "Estructura de la piel en seis especies de dragonet (Gobiesociformes; Callionymidae): diferencias interespecíficas en tipos de células glandulares y secreción de moco". Revista de biología de peces . 66 (5): 1411–1418. doi : 10.1111 / j.0022-1112.2005.00692.x .
- ^ ¿Los atunes tienen escamas? Centro de Ciencias Pesqueras del Noreste, NOAA Fisheries. Consultado el 4 de agosto de 2019.
- ^ Levítico 11: 9-10
- ^ a b c Aryeh Citron, "Todo sobre el pescado kosher"
- ^ Verificación de la certificación Kosher Fish OU . Consultado el 9 de agosto de 2019.
- ^ a b Lee, HJ; Kusche, H .; Meyer, A. (2012). "Comportamiento de forrajeo de manos en peces cíclidos que comen escamas: su papel potencial en la conformación de la asimetría morfológica" . PLOS ONE . 7 (9): e44670. Código Bibliográfico : 2012PLoSO ... 744670L . doi : 10.1371 / journal.pone.0044670 . PMC 3435272 . PMID 22970282 .
- ^ Froese, R. y D. Pauly. Editores. "Glosario: Lepidofagia" . FishBase . Consultado el 12 de abril de 2007 .
- ↑ a b c Janovetz, Jeff (2005). "Morfología funcional de la alimentación en el especialista en escamas Catoprion mento " (PDF) . La Revista de Biología Experimental . 208 (Pt 24): 4757–4768. doi : 10.1242 / jeb.01938 . PMID 16326957 . S2CID 15566769 .
- ^ Martín, C .; PC Wainwright (2011). "La novedad trófica está relacionada con tasas excepcionales de diversificación morfológica en dos radiaciones adaptativas de cachorrito Cyprinodon" . Evolución . 65 (8): 2197–2212. doi : 10.1111 / j.1558-5646.2011.01294.x . PMID 21790569 . S2CID 23695342 .
- ^ Hori, M. (1993). "Selección natural dependiente de la frecuencia en la mano de los peces cíclidos que comen escamas". Ciencia . 260 (5105): 216–219. Código Bibliográfico : 1993Sci ... 260..216H . doi : 10.1126 / science.260.5105.216 . PMID 17807183 . S2CID 33113282 .
- ^ Stewart, TA; Albertson, RC (2010). "Evolución de un aparato de alimentación depredador único: anatomía funcional, desarrollo y un locus genético para la lateralidad de la mandíbula en los cíclidos que comen escamas del lago Tanganica" . Biología BMC . 8 (1): 8. doi : 10.1186 / 1741-7007-8-8 . PMC 2828976 . PMID 20102595 .
Lectura adicional
- Helfman, GS, BB Collette y DE Facey (1997). La diversidad de peces . Ciencia Blackwell. págs. 33–36. ISBN 978-0-86542-256-8.CS1 maint: multiple names: authors list (link)
- Schultze, HP (2016). "Escamas, esmalte, cosmina, ganoína y osteictios tempranos" . Comptes Rendus Palevol . 15 (1–2): 83–102. doi : 10.1016 / j.crpv.2015.04.001 .
| vtmiPez | |||||
|---|---|---|---|---|---|
| About fish |
| ||||
| Anatomy andphysiology |
| ||||
| Sensorysystems |
| ||||
| Reproduction |
| ||||
| Locomotion |
| ||||
| Otherbehaviour |
| ||||
| By habitat |
| ||||
| Other types |
| ||||
| Commerce |
| ||||
| Major groups |
| ||||
| Lists |
| ||||
| |||||
Enlaces externos
- Medios de Wikimedia Commons
- Aspectos hidrodinámicos de las escamas de tiburón [1]
- Escamas de pescado y manipulación del flujo [2]