
El sesgo de GC es cuando los nucleótidos guanina y citosina son abundantes o insuficientemente abundantes en una región particular de ADN o ARN . En condiciones de equilibrio (sin presión mutacional o selectiva y con nucleótidos distribuidos aleatoriamente dentro del genoma ) hay una frecuencia igual de las cuatro bases de ADN ( adenina , guanina , timina y citosina ) en ambas cadenas simples de una molécula de ADN. [1] Sin embargo, en la mayoría de las bacterias (p . Ej., E. coli) y algunas arqueas (por ejemplo, Sulfolobus solfataricus ), las composiciones de nucleótidos son asimétricas entre la hebra principal y la hebra rezagada : la hebra principal contiene más guanina (G) y timina (T), mientras que la hebra rezagada contiene más adenina (A) y citosina (C). [1] Este fenómeno se conoce como sesgo de GC y AT . Se representa matemáticamente de la siguiente manera: [2]
Desviación de GC = (G - C) / (G + C)
En sesgo = (A - T) / (A + T)
Composición asimétrica de nucleótidos
El trabajo de Erwin Chargaff en 1950 demostró que, en el ADN, las bases guanina y citosina se encuentran en igual abundancia, y las bases adenina y timina se encuentran en igual abundancia. Sin embargo, no hubo igualdad entre la cantidad de un par frente al otro. [3] El hallazgo de Chargaff se conoce como regla de Chargaff o regla de paridad 1 . [3] Tres años más tarde, Watson y Crick utilizaron este hecho durante su derivación de la estructura del ADN, su modelo de doble hélice .
Un resultado natural de la regla de paridad 1, en el estado de equilibrio, en el que no hay sesgos de mutación y / o selección en ninguna de las dos hebras de ADN, es que cuando hay una tasa de sustitución igual, los nucleótidos complementarios de cada hebra tienen cantidades iguales de una base dada y su complemento. [4] En otras palabras, en cada cadena de ADN la frecuencia de aparición de T es igual a A y la frecuencia de aparición de G es igual a C porque la tasa de sustitución es presumiblemente igual. Este fenómeno se conoce como regla de paridad 2 . Por lo tanto, la segunda regla de paridad solo existe cuando no hay mutación o sustitución.
Cualquier desviación de la regla de paridad 2 dará como resultado una composición de base asimétrica que discrimina la hebra principal, es decir, la hebra de ADN que se replica en la dirección de avance, de la hebra rezagada. Esta asimetría se conoce como sesgo GC o AT. [1]
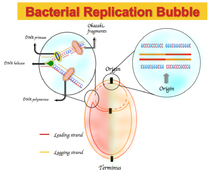
En algunos genomas bacterianos, hay un enriquecimiento de guanina sobre citosina y timina sobre adenina en la hebra principal y viceversa para la hebra rezagada. El espectro de sesgo de la composición de nucleótidos varía de -1, que corresponde a G = 0 o A = 0, a +1, que corresponde a T = 0 o C = 0. [1] Por lo tanto, el sesgo de GC positivo representa la riqueza de G sobre C y el sesgo de GC negativo representa la riqueza de C sobre G. Como resultado, se espera ver un sesgo de GC positivo y un sesgo de AT negativo en la cadena principal, y un sesgo de GC negativo y un sesgo de AT positivo en la cadena retrasada. [5] Los cambios de sesgo de GC o AT firman en los límites de los dos replichores , que corresponden al origen o al término de la replicación del ADN. [1] [4] [5]Originalmente, esta composición asimétrica de nucleótidos se explicó como un mecanismo diferente utilizado en la replicación del ADN entre la hebra principal y la hebra retrasada. La replicación del ADN es semi-conservadora y un proceso asimétrico en sí mismo. [6] Esta asimetría se debe a la formación de la bifurcación de replicación y su división en hebras iniciales y rezagadas nacientes. La cadena principal se sintetiza continuamente y en yuxtaposición a la cadena principal; la hebra rezagada se replica a través de fragmentos cortos de polinucleótido ( fragmentos de Okazaki ) en una dirección de 5 'a 3'. [6]
Gráficos de sesgo de cálculo y GC
Hay tres enfoques principales para calcular y demostrar gráficamente el sesgo de GC y sus propiedades.
Asimetría GC
El primer enfoque es la asimetría GC y AT. [1] Jean R. Lobry fue el primero en informar, en 1996, [7] la presencia de asimetría composicional en los genomas de tres bacterias: E. coli , Bacillus subtilis y Haemophilus influenzae . Las fórmulas originales en ese momento no se llamaban sesgo, sino desviación de [A] = [T] o [C] = [G]:
desviación de [A] = [T] como (A - T) / (A + T);
desviación de [C] = [G] como (C - G) / (C + G);
donde A, T, G y C representan la frecuencia de aparición de la base equivalente en una secuencia particular en una longitud definida. Se utiliza una estrategia de ventana deslizante para calcular la desviación de C a través del genoma. En estas gráficas, una desviación positiva de C corresponde a la hebra rezagada y una desviación negativa de C corresponde a la hebra principal. [8]Además, el lugar donde cambia la señal de desviación corresponde al origen o terminal. El eje x representa las ubicaciones de los cromosomas trazadas de 5 ′ a 3 ′ y el eje y representa el valor de desviación. La principal debilidad de este método es su propiedad dependiente del tamaño de la ventana. Por lo tanto, elegir un tamaño de ventana adecuado afecta en gran medida el resultado del gráfico. Otras técnicas deben combinarse con la desviación para identificar y localizar el origen de la replicación del ADN con mayor precisión.
Desviación CGC

El segundo enfoque se conoce como sesgo de GC acumulativo (sesgo de CGC). [2] Este método todavía utiliza la estrategia de ventana deslizante pero aprovecha la suma de las ventanas adyacentes desde un inicio arbitrario. En este esquema, el genoma completo generalmente se traza de 5 'a 3' usando un comienzo arbitrario y una hebra arbitraria. En el gráfico de sesgo de GC acumulativo, los picos corresponden a los puntos de conmutación (terminal u origen).
En contraste con el artículo anterior de Lobry, las implementaciones recientes de GC sesgo invierten la definición original, redefiniéndola para ser:
Desviación de GC = (G - C) / (G + C).
Con la definición invertida de sesgo de GC, el valor máximo del sesgo acumulativo corresponde al terminal y el valor mínimo corresponde al origen de la replicación.
Curva Z
La aproximación final es la curva Z . [9] A diferencia de los métodos anteriores, este método no utiliza la estrategia de ventana deslizante y se cree que funciona mejor para encontrar el origen de la replicación. [9] En este método, se investiga la frecuencia acumulada de cada base con respecto a la base al comienzo de la secuencia. La curva Z utiliza una representación tridimensional con los siguientes parámetros:



Donde , representa el exceso de purina sobre pirimidina, denota exceso de ceto sobre amino, y muestra la relación entre los enlaces de hidrógeno débiles y fuertes .y los componentes pueden detectar por sí solos el origen de replicación y la composición asimétrica de las hebras. Debería utilizarse una combinación de estos métodos para la predicción del origen y la terminal de la replicación, con el fin de compensar su debilidad.






Mecanismo
Existe una falta de consenso en la comunidad científica con respecto al mecanismo subyacente al sesgo en la composición de nucleótidos dentro de cada hebra de ADN. Hay dos escuelas de pensamiento principales que explican el mecanismo detrás de la composición de nucleótidos específicos de la cadena en las bacterias. [4]
El primero describe un sesgo y una presión mutacional asimétrica en cada hebra de ADN durante la replicación y la transcripción . [4] [10] Debido a la naturaleza asimétrica del proceso de replicación, una frecuencia de mutaciones desiguales y una eficiencia de reparación del ADN durante el proceso de replicación pueden introducir más mutaciones en una hebra en comparación con la otra. [5] Además, el tiempo utilizado para la replicación entre las dos hebras varía y puede conducir a una presión mutacional asimétrica entre la hebra principal y la rezagada. [11] Además de las mutaciones durante la replicación del ADN, las mutaciones transcripcionales pueden crear un sesgo en la composición de nucleótidos específica de la hebra.[5] La desaminación de la citosina y, en última instancia, la mutación de la citosina a timina en una hebra de ADN puede aumentar el número relativo de guanina y timina a citosina y adenina. [5] En la mayoría de las bacterias, la mayoría de los genes están codificados en la cadena principal. [4] Por ejemplo, la cadena principal de Bacillus subtilis codifica el 75% de los genes. [5] Además, se ha informado de un exceso de desaminación y conversión de citosina en timina en la hebra codificante en comparación con la hebra no codificante. [4] [5] [12] Una posible explicación es que la hebra no transcrita ( hebra codificante) es monocatenario durante el proceso de transcripción; por lo tanto, es más vulnerable a la desaminación en comparación con la hebra transcrita (hebra no codificante ). [5] [13] Otra explicación es que la actividad de reparación de desaminación durante la transcripción no ocurre en la cadena codificante. [5] Solo la hebra transcrita se beneficia de estos eventos de reparación de la desaminación.
La segunda escuela de pensamiento describe el mecanismo de la desviación de GC y AT como resultado de una diferencia en la presión selectiva entre las hebras principales y rezagadas. [4] [5] [13] El examen del genoma procariótico muestra una preferencia en la posición del tercer codón para G sobre C y T sobre A. [5] Esta discriminación crea una composición de nucleótidos asimétrica, si la hebra codificante está distribuida de manera desigual entre los hebras delanteras y rezagadas, como en el caso de las bacterias. Además, se ha demostrado que los genes altamente transcritos, como las proteínas ribosómicas , se encuentran principalmente en la cadena principal de las bacterias. [5]Por lo tanto, un sesgo en la elección del codón de tercera posición de G sobre C puede conducir a un sesgo de GC. Además, algunas secuencias de señales son ricas en guanina y timina, como las secuencias chi , y estas secuencias pueden tener una mayor frecuencia de aparición en una hebra en comparación con la otra. [4] [5]
Tanto la presión mutacional como la selectiva pueden introducir asimetría de forma independiente en las cadenas de ADN. Sin embargo, la combinación y el efecto acumulativo de ambos mecanismos es la explicación más plausible del sesgo de GC y AT. [4] [13]
Usos
Se ha demostrado que el sesgo de GC es útil como indicador de la hebra principal, hebra retrasada, origen de replicación y terminal de replicación del ADN. [1] [4] [5] La mayoría de las bacterias y arqueas contienen solo un origen de replicación del ADN. [1] El sesgo de GC es positivo y negativo en la cadena principal y en la cadena rezagada, respectivamente; por lo tanto, se espera ver un cambio en el signo de sesgo de GC justo en el punto de origen y terminación de la replicación del ADN. [4] El sesgo de GC también se puede utilizar para estudiar los sesgos de hebra y el mecanismo relacionado con ellos mediante el cálculo del exceso de una base sobre su base complementaria en diferentes medios. [4] [5] [13] Los métodos como el sesgo de GC, el sesgo de CGC y la curva Z son herramientas que pueden brindar la oportunidad de investigar mejor el mecanismo de replicación del ADN en diferentes organismos.
Referencias
- ^ a b c d e f g h Lobry, JR Patrones de sustitución asimétrica en las dos hebras de ADN de las bacterias. Biología molecular y evolución 13, 660-665 (1996).
- ↑ a b Grigoriev, A. Análisis de genomas con diagramas de sesgo acumulativo. Nucleic Acids Research 26, 2286-2290 (1998).
- ^ a b Chargaff, E. Especificidad química de los ácidos nucleicos y mecanismo de su degradación enzimática. Experientia 6, 201-209 (1950).
- ^ a b c d e f g h i j k l Necsulea, A. & Lobry, JR Un nuevo método para evaluar el efecto de la replicación en la asimetría de la composición de la base del ADN. Biología molecular y evolución 24, 2169-2179, doi: 10.1093 / molbev / msm148 (2007).
- ^ a b c d e f g h i j k l m n o Tillier, ER y Collins, RA Las contribuciones de la orientación de la replicación, la dirección de los genes y las secuencias de señales a las asimetrías de composición de bases en los genomas bacterianos. Journal of molecular evolution 50, 249-257 (2000).
- ^ a b Rocha, EP La organización relacionada con la replicación de los genomas bacterianos. Microbiology 150, 1609-1627, doi: 10.1099 / mic.0.26974-0 (2004).
- ^ Lobry, JR (mayo de 1996). "Patrones de sustitución asimétrica en las dos hebras de ADN de bacterias" . Biología Molecular y Evolución . 13 (5): 660–665. doi : 10.1093 / oxfordjournals.molbev.a025626 . ISSN 0737-4038 . PMID 8676740 .
- ^ "Explicación del artículo de Lobry 1996"
- ^ a b Zhang, R. & Zhang, CT Orígenes de replicación múltiple de la especie archaeon Halobacterium NRC-1. Comunicaciones de investigación bioquímica y biofísica 302, 728-734 (2003).
- ^ Lobry, JR & Sueoka, N. Presiones de mutación direccional asimétrica en bacterias. Biología del genoma 3, RESEARCH0058 (2002).
- ^ Eppinger, M., Baar, C., Raddatz, G., Huson, DH & Schuster, SC Análisis comparativo de cuatro Campylobacterales. Reseñas de la naturaleza. Microbiology 2, 872-885, doi: 10.1038 / nrmicro1024 (2004).
- ^ Marin, A. y Xia, X. GC sesgo en genes que codifican proteínas entre las cadenas principales y rezagadas en los genomas bacterianos: nuevos modelos de sustitución que incorporan el sesgo de la cadena. Revista de biología teórica 253, 508-513, doi: 10.1016 / j.jtbi.2008.04.004 (2008).
- ^ a b c d Charneski CA, Honti F., Bryant JM, Hurst LD, Feil EJ Atypical AT Skew in Firmicute Genomes Resultados de la selección y no de la mutación. PLoS Genetics 7 (9): e1002283 (2011).
- Mewes, HW y col. MIPS: análisis y anotación de proteínas de genomas completos en 2005. Nucleic Acids Res 34, D169-172, doi: 10.1093 / nar / gkj148 (2006).