| Clase MHC | |
|---|---|
 Representación esquemática de MHC clase I | |
| Identificadores | |
| Símbolo | MHC clase I |
| Membranome | 63 |
Las moléculas de MHC de clase I son una de las dos clases principales de moléculas del complejo principal de histocompatibilidad (MHC) (la otra es MHC de clase II ) y se encuentran en la superficie celular de todas las células nucleadas en los cuerpos de los vertebrados . [1] [2] También se presentan en las plaquetas , pero no en los glóbulos rojos . Su función es mostrar fragmentos de péptidos de proteínas desde el interior de la célula a las células T citotóxicas ; esto desencadenará una respuesta inmediata del sistema inmunológico contra un antígeno no propio particular que se muestra con la ayuda de una proteína MHC de clase I. Debido a que las moléculas de MHC de clase I están presentespéptidos derivados de proteínas citosólicas , la vía de presentación del MHC de clase I a menudo se denomina vía citosólica o endógena . [3]
En los seres humanos, los HLA correspondientes al MHC de clase I son HLA-A , HLA-B y HLA-C .
Función
Las moléculas de MHC de clase I se unen a péptidos generados principalmente a partir de la degradación de proteínas citosólicas por el proteasoma . El complejo MHC I: péptido se inserta a través del retículo endoplásmico en la membrana plasmática externa de la célula. El péptido epítopo se une a partes extracelulares de la molécula del MHC de clase I. Por tanto, la función del MHC de clase I es mostrar proteínas intracelulares a las células T citotóxicas (CTL). Sin embargo, el MHC de clase I también puede presentar péptidos generados a partir de proteínas exógenas, en un proceso conocido como presentación cruzada .
Una célula normal mostrará péptidos de la renovación de proteínas celulares normales en su MHC de clase I, y los CTL no se activarán en respuesta a ellos debido a los mecanismos de tolerancia centrales y periféricos. Cuando una célula expresa proteínas extrañas, como después de una infección viral, una fracción del MHC de clase I mostrará estos péptidos en la superficie celular. En consecuencia, los CTL específicos para el complejo MHC: péptido reconocerán y matarán las células presentadoras.
Alternativamente, el MHC de clase I en sí mismo puede servir como un ligando inhibidor para las células asesinas naturales (NK). La reducción de los niveles normales de MHC de clase I de superficie, un mecanismo empleado por algunos virus [4] y ciertos tumores para evadir las respuestas de los CTL, activa la muerte de las células NK.
PirB y plasticidad visual
El receptor B pareado de tipo inmunoglobulina (PirB), un receptor de unión a MHCI, participa en la regulación de la plasticidad visual . [5] PirB se expresa en el sistema nervioso central y disminuye la plasticidad de dominancia ocular en el período crítico del desarrollo y la edad adulta. [5] Cuando se abolió la función de PirB en ratones mutantes, la plasticidad de dominancia ocular se hizo más pronunciada en todas las edades. [5] Los ratones mutantes con pérdida de función de PirB también exhibieron una mayor plasticidad después de la privación monocular durante el período crítico . [5]Estos resultados sugieren que PirB puede estar involucrado en la modulación de la plasticidad sináptica en la corteza visual .
Estructura
Las moléculas de MHC de clase I son heterodímeros que constan de dos cadenas polipeptídicas, α y β 2 -microglobulina (B2M). Las dos cadenas están unidas de forma no covalente a través de interacción de B2M y la α 3 dominio. Solo la cadena α es polimórfica y está codificada por un gen HLA , mientras que la subunidad B2M no es polimórfica y está codificada por el gen de la microglobulina Beta-2 . La α 3 dominio es que abarca la membrana plasmática e interactúa con el CD8 co-receptor de células T . La interacción α 3 -CD8 mantiene la molécula MHC I en su lugar mientras que el receptor de células T (TCR) en la superficie de la célula T citotóxica se une a su α 1 -α 2ligando heterodímero y comprueba la antigenicidad del péptido acoplado. Los dominios α 1 y α 2 se pliegan para formar un surco para que los péptidos se unan. Las moléculas de MHC de clase I se unen a péptidos que tienen predominantemente de 8 a 10 aminoácidos de longitud (Parham 87), pero también se ha informado de la unión de péptidos más largos. [6]
Síntesis
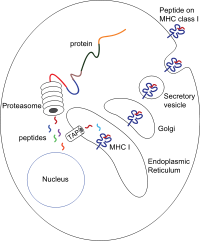
Los péptidos son generados principalmente en el citosol por el proteasoma . El proteasoma es una macromolécula que consta de 28 subunidades, de las cuales la mitad afecta la actividad proteolítica . El proteasoma degrada las proteínas intracelulares en pequeños péptidos que luego se liberan en el citosol. Los proteasomas también pueden ligar distintos fragmentos de péptidos (denominados péptidos empalmados), produciendo secuencias que no son contiguas y, por lo tanto, no tienen una plantilla lineal en el genoma. El origen de los segmentos de péptidos empalmados puede ser de la misma proteína (empalme en cis) o de proteínas diferentes (empalme en trans). [7] [8] Los péptidos deben trasladarse del citosol al retículo endoplásmico.(ER) para encontrar la molécula del MHC de clase I, cuyo sitio de unión al péptido se encuentra en la luz del ER. Tienen un pliegue de Ig proximal a la membrana
Translocación y carga de péptidos
La translocación del péptido desde el citosol al lumen del RE se realiza mediante el transportador asociado con el procesamiento de antígenos (TAP). TAP es un miembro de la familia de transportadores ABC y es un polipéptido heterodimérico que abarca múltiples membranas que consta de TAP1 y TAP2. Las dos subunidades forman un sitio de unión de péptidos y dos sitios de unión de ATP que se enfrentan al citosol. TAP se une a los péptidos en el lado citoplásmico y los transloca bajo consumo de ATP en la luz del RE. La molécula del MHC de clase I, a su vez, se carga con péptidos en la luz del RE.
El proceso de carga de péptidos involucra varias otras moléculas que forman un gran complejo multimérico llamado complejo de carga de péptidos [9] que consiste en TAP, tapasina , calreticulina , calnexina y Erp57 ( PDIA3 ). La calnexina actúa para estabilizar las cadenas α del MHC de clase I antes de la unión de β2m. Después del ensamblaje completo de la molécula de MHC, la calnexina se disocia. La molécula de MHC que carece de un péptido unido es inherentemente inestable y requiere la unión de las chaperonas calreticulina y Erp57. Además, la tapasina se une a la molécula del MHC y sirve para unirla a las proteínas TAP y facilita la selección de péptidos en un proceso iterativo llamado edición de péptidos [10].[11] [12] facilitando así una mayor carga de péptidos y colocalización.
Una vez que el péptido se carga en la molécula del MHC de clase I, el complejo se disocia y sale del RE a través de la vía secretora para llegar a la superficie celular. El transporte de las moléculas del MHC de clase I a través de la vía secretora implica varias modificaciones postraduccionales de la molécula del MHC. Algunas de las modificaciones postraduccionales ocurren en el RE e implican cambios en las regiones de N-glicanos de la proteína, seguidas de cambios extensos en los N-glicanos en el aparato de Golgi . Los N-glicanos maduran completamente antes de llegar a la superficie celular.
Eliminación de péptidos
Los péptidos que no se unen a las moléculas del MHC de clase I en la luz del retículo endoplásmico (RE) se eliminan del RE a través del canal sec61 hacia el citosol, [13] [14] donde podrían sufrir un mayor recorte de tamaño y podrían ser translocado por TAP de nuevo en ER para unirse a una molécula de MHC de clase I.
Por ejemplo, se ha observado una interacción de sec61 con albúmina bovina . [15]
Efecto de los virus
Las moléculas de MHC de clase I están cargadas con péptidos generados a partir de la degradación de proteínas citosólicas ubiquitinadas en proteasomas . A medida que los virus inducen la expresión celular de proteínas virales, algunos de estos productos se marcan para su degradación, y los fragmentos de péptidos resultantes entran en el retículo endoplásmico y se unen a las moléculas del MHC I. De esta forma, la vía de presentación de antígenos dependiente del MHC de clase I, las células infectadas por el virus señalan a las células T que se están produciendo proteínas anormales como resultado de la infección.
El destino de la célula infectada por el virus es casi siempre la inducción de apoptosis a través de la inmunidad mediada por células , lo que reduce el riesgo de infectar células vecinas. Como respuesta evolutiva a este método de vigilancia inmunitaria, muchos virus pueden regular a la baja o prevenir de otro modo la presentación de moléculas MHC de clase I en la superficie celular. A diferencia de los linfocitos T citotóxicos, el asesino naturalLas células (NK) normalmente se inactivan al reconocer moléculas de MHC I en la superficie de las células. Por lo tanto, en ausencia de moléculas MHC I, las células NK se activan y reconocen la célula como aberrante, lo que sugiere que puede estar infectada por virus que intentan evadir la destrucción inmunológica. Varios cánceres humanos también muestran una regulación a la baja de MHC I, dando a las células transformadas la misma ventaja de supervivencia de poder evitar la vigilancia inmunológica normal diseñada para destruir cualquier célula infectada o transformada. [dieciséis]
Genes e isotipos
- Muy polimórfico
- HLA-A ( HLA-A )
- HLA-B ( HLA-B )
- HLA-C ( HLA-C )
- Menos polimórfico
- HLA-E ( HLA-E )
- HLA-F ( HLA-F )
- HLA-G ( HLA-G )
- HLA-K (pseudogén)
- HLA-L (pseudogén)
Historia evolutiva
Los genes MHC de clase I se originaron en el ancestro común más reciente de todos los vertebrados con mandíbula y se han encontrado en todos los vertebrados con mandíbula vivos que se han estudiado hasta ahora. [2] Desde su aparición en vertebrados con mandíbulas, esta familia de genes ha estado sujeta a muchos caminos evolutivos divergentes a medida que se han producido eventos de especiación . Sin embargo, hay casos documentados de polimorfismos de especies trans en los genes del MHC de clase I, donde un alelo particular en un gen del MHC de clase I relacionado con la evolución permanece en dos especies, probablemente debido a una fuerte selección de equilibrio mediada por patógenos por patógenos que pueden infectar a ambos. especies.[17] La evolución del nacimiento y la muerte es una de las explicaciones mecanicistas del tamaño de la familia de genes del MHC de clase I.
Nacimiento y muerte de genes MHC de clase I
La evolución del nacimiento y la muerte afirma que los eventos de duplicación de genes hacen que el genoma contenga múltiples copias de un gen que luego pueden experimentar procesos evolutivos separados. A veces, estos procesos dan como resultado la pseudogeneización (muerte) de una copia del gen, aunque a veces este proceso da como resultado dos genes nuevos con funciones divergentes. [18] Es probable que los loci humanos del MHC de clase Ib (HLA-E, -F y -G) así como los pseudogenes del MHC de clase I surgieran de los loci del MHC de clase Ia (HLA-A, -B y -C) en este proceso de nacimiento y muerte. [19]
Referencias
- ^ Hewitt EW (octubre de 2003). "La vía de presentación del antígeno MHC de clase I: estrategias para la evasión inmune viral" . Inmunologia . 110 (2): 163–9. doi : 10.1046 / j.1365-2567.2003.01738.x . PMC 1783040 . PMID 14511229 .
- ↑ a b Kulski JK, Shiina T, Anzai T, Kohara S, Inoko H (diciembre de 2002). "Análisis genómico comparativo del MHC: la evolución de los bloques de duplicación de clase I, diversidad y complejidad del tiburón al hombre". Revisiones inmunológicas . 190 : 95-122. doi : 10.1034 / j.1600-065x.2002.19008.x . PMID 12493009 .
- ^ http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/H/HLA.html#Class_I_Histocompatibility_Molecules Páginas de biología de Kimball, moléculas de histocompatibilidad
- ^ Hansen TH, Bouvier M (julio de 2009). "Presentación del antígeno MHC clase I: aprender de las estrategias de evasión viral". Reseñas de la naturaleza. Inmunologia . 9 (7): 503-13. doi : 10.1038 / nri2575 . PMID 19498380 .
- ↑ a b c d Syken J, Grandpre T, Kanold PO, Shatz CJ (septiembre de 2006). "PirB restringe la plasticidad de dominancia ocular en la corteza visual". Ciencia . 313 (5794): 1795–800. Código bibliográfico : 2006Sci ... 313.1795S . doi : 10.1126 / science.1128232 . PMID 16917027 .
- ^ Burrows SR, Rossjohn J, McCluskey J (enero de 2006). "¿Nos hemos acortado demasiado en el mapeo de epítopos CTL?". Tendencias en inmunología . 27 (1): 11–6. doi : 10.1016 / j.it.2005.11.001 . PMID 16297661 .
- ^ Faridi, Pouya; Li, Chen; Ramarathinam, Sri H .; Vivian, Julian P .; Illing, Patricia T .; Mifsud, Nicole A .; Ayala, Rochelle; Song, Jiangning; Gearing, Linden J .; Hertzog, Paul J .; Ternette, Nicola; Rossjohn, Jamie; Croft, Nathan P .; Purcell, Anthony W. (12 de octubre de 2018). "Un subconjunto de péptidos HLA-I no tiene plantilla genómica: evidencia de ligandos de péptidos empalmados en cis y trans" (PDF) . Inmunología de la ciencia . 3 (28): eaar3947. doi : 10.1126 / sciimmunol.aar3947 . PMID 30315122 .
- ^ Liepe, Juliane; Marino, Fabio; Sidney, John; Jeko, Anita; Bunting, Daniel E .; Sette, Alessandro; Kloetzel, Peter M .; Stumpf, Michael PH; Diablos, Albert JR; Mishto, Michele (21 de octubre de 2016). "Una gran fracción de ligandos HLA de clase I son péptidos empalmados generados por proteasomas" (PDF) . Ciencia . 354 (6310): 354–358. Código bibliográfico : 2016Sci ... 354..354L . doi : 10.1126 / science.aaf4384 . hdl : 10044/1/42330 . PMID 27846572 .
- ^ Sangra A, Januliene D, Hofmann T, Koller N, Schmidt C, Trowitzsch S, Moeller A, Tampé R (noviembre de 2017). "Estructura del complejo de carga de péptido MHC-I humano". Naturaleza . 551 (7681): 525–528. Código Bib : 2017Natur.551..525B . doi : 10.1038 / nature24627 . PMID 29107940 .
- ^ Howarth M, Williams A, Tolstrup AB, Elliott T (agosto de 2004). "Tapasin mejora la presentación del péptido MHC de clase I según la vida media del péptido" . Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 101 (32): 11737–42. Código Bibliográfico : 2004PNAS..10111737H . doi : 10.1073 / pnas.0306294101 . PMC 511045 . PMID 15286279 .
- ^ Wearsch PA, Cresswell P (agosto de 2007). "Carga selectiva de péptidos de alta afinidad en moléculas de clase I del complejo principal de histocompatibilidad por el heterodímero tapasin-ERp57". Inmunología de la naturaleza . 8 (8): 873–81. doi : 10.1038 / ni1485 . PMID 17603487 .
- ^ Thirdborough SM, Roddick JS, Radcliffe JN, Howarth M, Stevenson FK, Elliott T (febrero de 2008). "Tapasin da forma a las jerarquías de inmunodominancia según la estabilidad cinética de los complejos péptido-MHC clase I". Revista europea de inmunología . 38 (2): 364–9. doi : 10.1002 / eji.200737832 . PMID 18196518 .
- ^ Koopmann JO, Albring J, Hüter E, Bulbuc N, Spee P, Neefjes J, Hämmerling GJ, Momburg F, et al. (Julio de 2000). "La exportación de péptidos antigénicos del retículo endoplásmico se cruza con la translocación de proteínas retrógradas a través del canal Sec61p" . La inmunidad . 13 (1): 117–27. doi : 10.1016 / S1074-7613 (00) 00013-3 . PMID 10933400 .
- ^ Albring J, Koopmann JO, Hämmerling GJ, Momburg F (enero de 2004). "La retrotranslocación de la cadena pesada del MHC de clase I del retículo endoplásmico al citosol depende del suministro de ATP al lumen del RE". Inmunología molecular . 40 (10): 733–41. doi : 10.1016 / j.molimm.2003.08.008 . PMID 14644099 .
- ^ Imai J, Hasegawa H, Maruya M, Koyasu S, Yahara I (enero de 2005). "Los antígenos exógenos se procesan a través de la degradación asociada al retículo endoplásmico (ERAD) en presentación cruzada por células dendríticas" . Inmunología internacional . 17 (1): 45–53. doi : 10.1093 / intimm / dxh184 . PMID 15546887 .
- ^ Wang Z, Zhang L, Qiao A, Watson K, Zhang J, Fan GH (febrero de 2008). "La activación de CXCR4 desencadena la ubiquitinación y la regulación a la baja del complejo principal de histocompatibilidad clase I (MHC-I) en las células HeLa del carcinoma epitelioide" . La revista de química biológica . 283 (7): 3951–9. doi : 10.1074 / jbc.m706848200 . PMID 18083706 .
- ^ Azevedo L, Serrano C, Amorim A, Cooper DN (septiembre de 2015). "El polimorfismo transespecífico en humanos y los grandes simios generalmente se mantiene equilibrando la selección que modula la respuesta inmune del huésped" . Genómica humana . 9 : 21. Doi : 10.1186 / s40246-015-0043-1 . PMC 4559023 . PMID 26337052 .
- ↑ Nei M, Rooney AP (14 de noviembre de 2005). "Evolución concertada y de nacimiento y muerte de familias multigénicas" . Revisión anual de genética . 39 (1): 121–52. doi : 10.1146 / annurev.genet.39.073003.112240 . PMC 1464479 . PMID 16285855 .
- ^ Hughes AL (marzo de 1995). "Origen y evolución de los pseudogenes HLA clase I" . Biología Molecular y Evolución . 12 (2): 247–58. doi : 10.1093 / oxfordjournals.molbev.a040201 . PMID 7700152 .
Enlaces externos
- Histocompatibilidad + Antígenos + Clase + I en los encabezados de temas médicos (MeSH) de la Biblioteca Nacional de Medicina de EE. UU .
- MHC + Class + I + Genes en los encabezados de materias médicas (MeSH) de la Biblioteca Nacional de Medicina de EE. UU .