| Parte de una serie sobre |
| Biología evolucionaria |
|---|
 |
|
En biología evolutiva , los paisajes de fitness o los paisajes adaptativos (tipos de paisajes evolutivos ) se utilizan para visualizar la relación entre los genotipos y el éxito reproductivo . Se supone que cada genotipo tiene una tasa de replicación bien definida (a menudo denominada aptitud). Esta aptitud es la "altura" del paisaje. Se dice que los genotipos que son similares están "cercanos" entre sí, mientras que los que son muy diferentes están "lejos" entre sí. El conjunto de todos los genotipos posibles, su grado de similitud y sus valores de aptitud relacionados se denomina paisaje de aptitud. La idea de un paisaje de aptitud es una metáfora para ayudar a explicar las formas defectuosas en la evolución por selección natural , incluidas las hazañas y los fallos en los animales, como sus reacciones a los estímulos sobrenaturales .
La idea de estudiar la evolución visualizando la distribución de los valores de aptitud como una especie de paisaje fue introducida por primera vez por Sewall Wright en 1932. [1]
En los problemas de optimización evolutiva , los paisajes de aptitud son evaluaciones de una función de aptitud para todas las soluciones candidatas (ver más abajo).
En biología

En todos los paisajes del fitness, la altura representa y es una metáfora visual del fitness . Hay tres formas distintas de caracterizar las otras dimensiones, aunque en cada caso la distancia representa y es una metáfora del grado de disimilitud. [2]
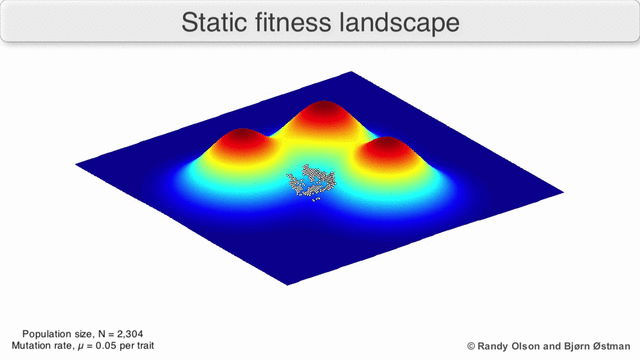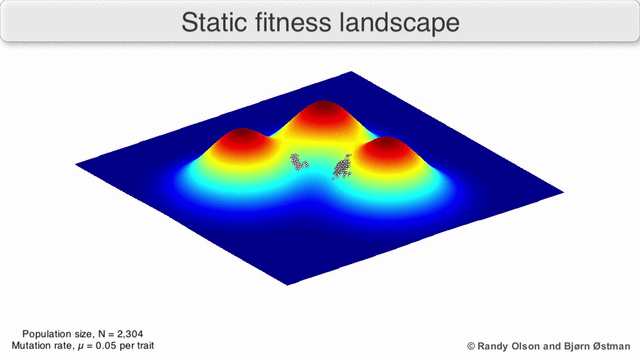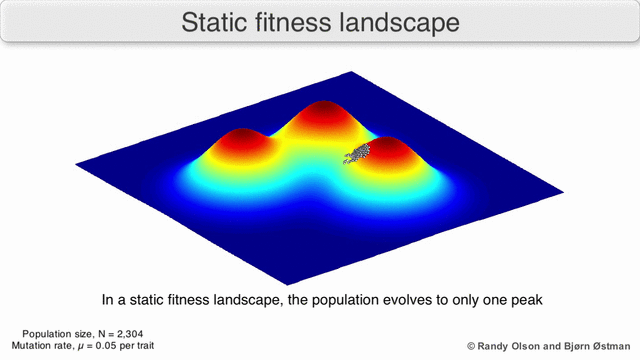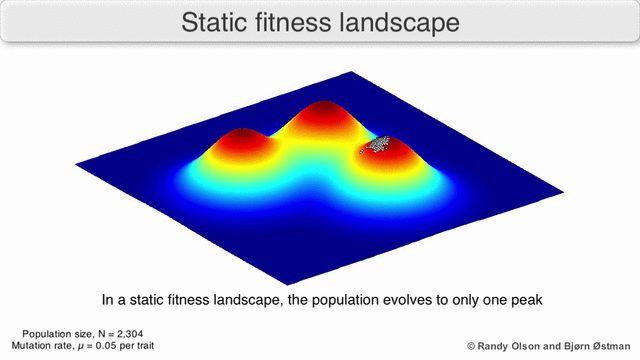
Los paisajes de fitness a menudo se conciben como cadenas montañosas. Existen picos locales (puntos desde los cuales todos los caminos son cuesta abajo, es decir, con menor aptitud) y valles (regiones desde las que muchos caminos conducen cuesta arriba). Un paisaje de fitness con muchos picos locales rodeados de valles profundos se llama accidentado. Si todos los genotipos tienen la misma tasa de replicación, por otro lado, se dice que un panorama de aptitud es plano. Una población en evolución generalmente sube cuesta arriba en el panorama de la aptitud, mediante una serie de pequeños cambios genéticos, hasta que, en el límite de tiempo infinito, se alcanza un óptimo local.
Tenga en cuenta que no siempre se puede encontrar un óptimo local, incluso en el tiempo evolutivo: si el óptimo local se puede encontrar en un período de tiempo razonable, entonces el panorama de aptitud se llama "fácil" y si el tiempo requerido es exponencial, entonces el panorama de aptitud se llama " duro". [3] Los paisajes duros se caracterizan por la propiedad de laberinto por la cual un alelo que alguna vez fue beneficioso se vuelve perjudicial, lo que obliga a la evolución a retroceder. Sin embargo, la presencia de la propiedad en forma de laberinto en paisajes de fitness inspirados biofísicamente puede no ser suficiente para generar un paisaje duro. [4]
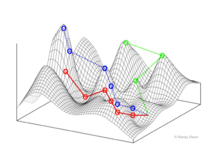
Genotipo para paisajes de fitness
Wright visualizó un espacio genotípico como un hipercubo . [1] No se define una "dimensión" continua del genotipo. En cambio, una red de genotipos está conectada a través de rutas mutacionales.
El modelo NK de Stuart Kauffman entra en esta categoría de paisaje de fitness. Las nuevas técnicas de análisis de redes, como los gráficos de atracción ponderados por selección (SWAG), también utilizan un espacio de genotipo adimensional. [5]
Frecuencia alélica de los paisajes de fitness
El trabajo matemático de Wright describió la aptitud como una función de las frecuencias alélicas. [2] Aquí, cada dimensión describe una frecuencia alélica en un gen diferente, y va entre 0 y 1.
Fenotipo para paisajes de fitness
En el tercer tipo de paisaje de fitness, cada dimensión representa un rasgo fenotípico diferente. [2] Bajo los supuestos de la genética cuantitativa, estas dimensiones fenotípicas pueden mapearse en genotipos. Consulte las visualizaciones a continuación para ver ejemplos de fenotipos de paisajes de fitness.
En optimización evolutiva
Aparte del campo de la biología evolutiva, el concepto de paisaje de fitness también ha ganado importancia en los métodos de optimización evolutiva , como los algoritmos genéticos o las estrategias de evolución . En la optimización evolutiva, uno intenta resolver problemas del mundo real (por ejemplo, problemas de ingeniería o logísticos ) imitando la dinámica de la evolución biológica. Por ejemplo, un camión de reparto con varias direcciones de destino puede tomar una gran variedad de rutas diferentes, pero solo unas pocas darán como resultado un tiempo de conducción corto.
Para utilizar muchas formas comunes de optimización evolutiva, uno tiene que definir para cada posible solución s al problema de interés (es decir, cada ruta posible en el caso del camión de reparto) cómo 'bueno' que es. Esto se realiza mediante la introducción de un escalar -valued función F (s) (valor escalar medios que f (s) es un número sencillo, tales como 0.3, mientras que s puede ser un objeto más complicado, por ejemplo, una lista de direcciones de destino en la caso del camión de reparto), que se denomina función de aptitud .
Una f (s) alta implica que s es una buena solución. En el caso del camión de reparto, f (s) podría ser el número de entregas por hora en la ruta s . La mejor solución, o al menos una muy buena, se encuentra de la siguiente manera: inicialmente, se crea una población de soluciones aleatorias. Luego, las soluciones se modifican y se seleccionan para aquellas con mayor aptitud, hasta que se encuentra una solución satisfactoria.
Las técnicas de optimización evolutiva son particularmente útiles en situaciones en las que es fácil determinar la calidad de una única solución, pero es difícil analizar todas las soluciones posibles una por una (es fácil determinar el tiempo de conducción para una ruta particular del camión de reparto). , pero es casi imposible verificar todas las rutas posibles una vez que el número de destinos aumenta a más de un puñado).
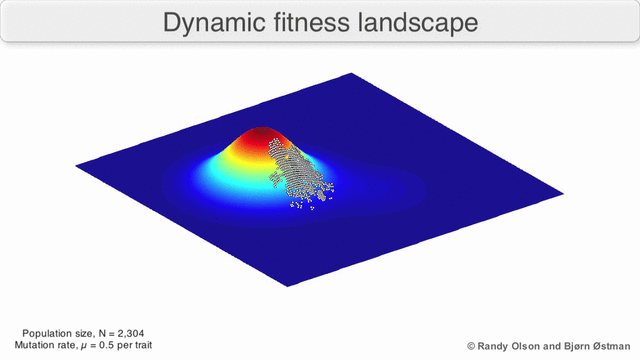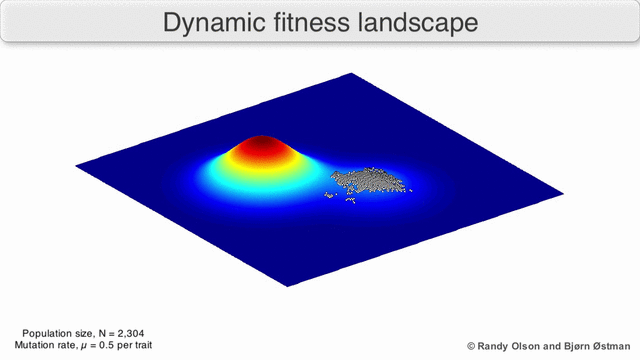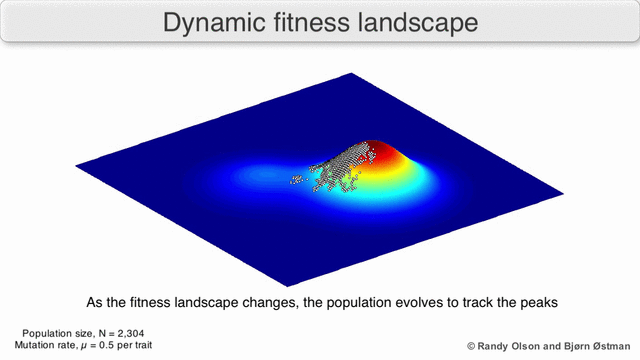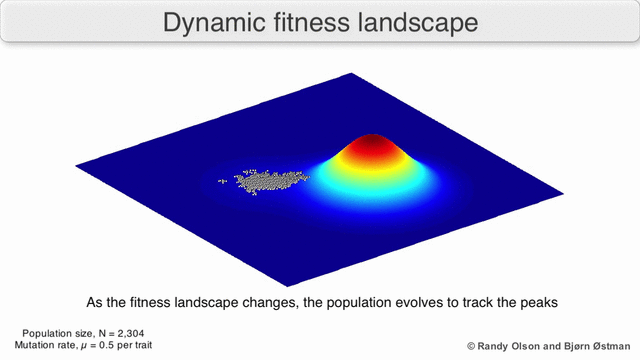
Incluso en los casos en los que es difícil definir una función de aptitud, el concepto de paisaje de aptitud puede resultar útil. Por ejemplo, si la evaluación de la aptitud se realiza mediante muestreo estocástico, el muestreo se realiza a partir de una distribución (generalmente desconocida) en cada punto; No obstante, puede ser útil razonar sobre el paisaje formado por la adecuación esperada en cada punto. Si la aptitud cambia con el tiempo (optimización dinámica) o con otras especies en el medio ambiente (coevolución), aún puede ser útil razonar sobre las trayectorias del panorama de aptitud instantánea. Sin embargo, en algunos casos (por ejemplo, computación evolutiva interactiva basada en preferencias) la relevancia es más limitada, porque no hay garantía de que las preferencias humanas sean consistentes con una única asignación de aptitud.
El concepto de función de aptitud f (s) con valor escalar también corresponde al concepto de función potencial o de energía en física . Los dos conceptos solo difieren en que los físicos tradicionalmente piensan en términos de minimizar la función potencial, mientras que los biólogos prefieren la noción de que se maximiza la aptitud. Por lo tanto, tomar la inversa de una función potencial la convierte en una función de aptitud, y viceversa [ cita requerida ] .
Advertencias y limitaciones
Existen varias salvedades importantes. Dado que la mente humana lucha por pensar en más de tres dimensiones, las topologías 3D pueden inducir a error cuando se habla de paisajes de fitness altamente multidimensionales. [6] [7] En particular, no está claro si los picos en los paisajes de aptitud biológica natural están realmente separados por valles de aptitud en paisajes tan multidimensionales, o si están conectados por crestas neutrales enormemente largas. [8] [9] Además, el panorama de la aptitud no es estático en el tiempo, sino que depende del entorno cambiante y la evolución de otros genes. [5] Por lo tanto, es más un paisaje marino, [10]afectando aún más la separación de los picos adaptativos. Además, es relevante tener en cuenta que un paisaje en general no es una función absoluta sino relativa. [11] Finalmente, dado que es común usar la función como un proxy para la aptitud cuando se habla de enzimas, cualquier actividad promiscua existe como paisajes superpuestos que juntos determinarán la aptitud final del organismo, lo que implica una brecha entre diferentes paisajes relativos coexistentes. [12]
Con estas limitaciones en mente, los paisajes de fitness pueden seguir siendo una forma instructiva de pensar sobre la evolución. Es fundamentalmente posible medir (aunque no visualizar) algunos de los parámetros de rugosidad del paisaje y de número de picos, altura, separación y agrupamiento. Los paisajes en 3D simplificados se pueden utilizar entre sí para representar visualmente las características relevantes. Además, los paisajes de aptitud de pequeños subconjuntos de vías evolutivas pueden construirse y visualizarse experimentalmente, lo que podría revelar características como picos y valles de aptitud. [5] Los paisajes de aptitud de las vías evolutivas indican los probables pasos evolutivos y puntos finales entre conjuntos de mutaciones individuales.
Ver también
- Aproximación de aptitud
- Función fitness
- Evolución
- Algoritmo genético
- Hábitat (ecología)
- Montañismo
- Modelo NK
- Función potencial
- Criticidad autoorganizada
- Teleología en biología
Referencias
- ↑ a b Wright, Sewall (1932). "Los roles de la mutación, la endogamia, el mestizaje y la selección en la evolución" (PDF) . Actas del Sexto Congreso Internacional de Genética . 1 (8): 355–66.
- ↑ a b c Provine, William B. (1986). Sewall Wright y biología evolutiva . Prensa de la Universidad de Chicago.[ página necesaria ]
- ^ Kaznatcheev, Artem (2019). "Complejidad computacional como una restricción última de la evolución" . Genética . 212 (1): 245–265. doi : 10.1534 / genetics.119.302000 . PMC 6499524 . PMID 30833289 .
- ^ Bertram, Jason; Masel, Joanna (abril de 2020). "La evolución optimiza rápidamente la estabilidad y la agregación en proteínas de celosía a pesar de los valles y laberintos del paisaje omnipresente" . Genética . 214 (4): 1047–1057. doi : 10.1534 / genetics.120.302815 . PMC 7153934 . PMID 32107278 .
- ^ a b c Steinberg, B; Ostermeier, M (2016). "Los cambios ambientales puentean los valles evolutivos" . Avances científicos . 2 (1): e1500921. Código bibliográfico : 2016SciA .... 2E0921S . doi : 10.1126 / sciadv.1500921 . PMC 4737206 . PMID 26844293 .
- ^ McCandlish, David M (2011). "Visualización de paisajes de fitness" . Evolución . 65 (6): 1544–58. doi : 10.1111 / j.1558-5646.2011.01236.x . PMC 3668694 . PMID 21644947 .
- ^ McGhee, George R. (2006). La geometría de la evolución: paisajes adaptativos y morfoespacios teóricos . ISBN 978-1-139-45995-2.[ página necesaria ]
- ^ Gavrilets, S. (2004). Paisajes de fitness y el origen de las especies . Prensa de la Universidad de Princeton. ISBN 978-0-691-11983-0.[ página necesaria ]
- ^ Kaplan, Jonathan (2008). "¿El fin de la metáfora del paisaje adaptativo?". Biología y Filosofía . 23 (5): 625–38. doi : 10.1007 / s10539-008-9116-z . S2CID 170649453 .
- ^ Mustonen, Ville; Lässig, Michael (2009). "De los paisajes de fitness a los paisajes marinos: dinámica de no equilibrio de selección y adaptación". Tendencias en Genética . 25 (3): 111–9. doi : 10.1016 / j.tig.2009.01.002 . PMID 19232770 .
- ↑ Woodcock, Glenn; Higgs, Paul G (1996). "Evolución de la población en un paisaje de fitness de pico único multiplicativo". Revista de Biología Teórica . 179 (1): 61–73. doi : 10.1006 / jtbi.1996.0049 . PMID 8733432 .
- ^ Díaz Ochoa, Juan G (2017). "Mecanismos elásticos multiescala: computación y evolución biológica". Revista de evolución molecular . 86 (1): 47–57. Código bibliográfico : 2018JMolE..86 ... 47D . doi : 10.1007 / s00239-017-9823-7 . PMID 29248946 . S2CID 22624633 .
Enlaces externos
- Ejemplos de paisajes de fitness visualizados
- Vídeo: Uso de paisajes de fitness para visualizar la evolución en acción
- Blog de BEACON: Evolución 101: paisajes de fitness
- Blog de pleiotropía: una discusión interesante de las contribuciones de Sergey Gavrilets
- Evolución del pez cachorro — UC Davis
- Evolución 101: teoría del equilibrio cambiante (figura al final de la página)
- Superposición de trayectorias evolutivas a paisajes de fitness en realidad virtual
- Otras lecturas
- Contrapeso: la evolución como movimiento a través de un paisaje de fitness: una discusión interesante (aunque defectuosa) sobre la evolución y los paisajes de fitness
- Ejemplo del uso de Paisajes Evolutivos para pensar y hablar sobre la evolución
- Hendrik Richter; Andries P. Engelbrecht (2014). Avances recientes en la teoría y aplicación de paisajes de fitness . ISBN 978-3-642-41888-4.
- Beerenwinkel, Niko; Pachter, Lior; Sturmfels, Bernd (2007). "Epistasis y formas de paisajes fitness". Statistica Sinica . 17 (4): 1317–42. arXiv : q-bio.PE/0603034 . Bibcode : 2006q.bio ..... 3034B . Señor 2398598 .
- Richard Dawkins (1996). Escalando el Monte Improbable . ISBN 0-393-03930-7.
- Sergey Gavrilets (2004). Paisajes de fitness y origen de especies . ISBN 978-0-691-11983-0.
- Stuart Kauffman (1995). En casa en el universo: la búsqueda de leyes de autoorganización y complejidad . ISBN 978-0-19-511130-9.
- Melanie Mitchell (1996). Introducción a los algoritmos genéticos (PDF) . ISBN 978-0-262-63185-3.
- Langdon, WB; Poli, R. (2002). "Capítulo 2 Paisajes de fitness" . Fundamentos de la programación genética . ISBN 3-540-42451-2.
- Stuart Kauffman (1993). Los orígenes del orden . ISBN 978-0-19-507951-7.
- Poelwijk, Frank J; Kiviet, Daniel J; Weinreich, Daniel M; Bronceados, Sander J (2007). "Los paisajes de aptitud empírica revelan caminos evolutivos accesibles". Naturaleza . 445 (7126): 383–6. Código Bibliográfico : 2007Natur.445..383P . doi : 10.1038 / nature05451 . PMID 17251971 . S2CID 4415468 .